© Catherine Chaput, 2020
Effet d'un niveau élevé de bêta-hydroxybutyrate (BHB)
au jour 45 post-partum sur la qualité transcriptomique et
épigénétique des embryons
Mémoire
Catherine Chaput
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Effet d’un niveau élevé de bêta-hydroxybutyrate
(BHB) au jour 45 post-partum sur la qualité
transcriptomique et épigénétique des embryons
Mémoire
Catherine Chaput
Sous la direction de :
ii Résumé
En début de lactation, la vache subit un stress important occasionné par l’impossibilité de combler l’ensemble de ses besoins énergétiques par sa consommation exogène. Cette période spécifique se caractérise par une balance énergétique négative, entraîne une utilisation excessive des réserves corporelles de l’animal et représente un défi métabolique important. Ironiquement, depuis maintenant plus de 40 ans, le système incite les producteurs laitiers à effectuer l’insémination au jour 60 post-partum, c’est-à-dire au moment où la vache rencontre un déficit métabolique. Ce déficit au moment de la conception aurait un impact chez la progéniture, notamment au niveau épigénétique. Ce projet consiste à documenter l’effet de la balance énergétique négative sur la qualité de l’embryon et, en l’occurrence, à proposer des pistes afin d’améliorer la fertilité des bovins laitiers.
La mesure du bêta-hydroxybutyrate (BHB) a été effectuée à partir d’échantillons sanguins entre 45 et 60 jours post-partum sur dix-huit vaches de race Holstein. Selon la mesure obtenue, la vache fut classée comme étant faible ou élevée en BHB, afin d’avoir au moins six vaches par groupe. Après un processus de stimulation ovarien, chaque vache fut inséminée et les embryons, récoltés. Pour chaque vache, deux embryons ont été transférés dans deux primipares, afin de déterminer subséquemment la persistance des marqueurs dans le matériel biologique. Grâce à la plate-forme EmbryoGENE, il fut possible de déterminer l’expression génique ainsi que l’état de méthylation de l’ADN des embryons récoltés.
Les résultats obtenus soutiennent l’existence d’une altération du métabolisme énergétique au niveau embryonnaire, notamment par la modification de la voie de signalisation de mTOR ainsi que celle des sirtuines. Cette altération semble se traduire par une dysfonction mitochondriale et une inhibition de la transcription, entraînant un freinage au niveau cellulaire, probablement dû à la programmation de l’embryon à utiliser ses réserves lipidiques lors de conditions importantes de stress.
iii Abstract
In early lactation, the cow undergoes an important stress generated by the impossibility of filling its entire energetic needs by exogenous consumption. This is characterized by a negative energy balance, excessive use of animal body reserves and represents an important metabolic challenge. Ironically, for more than 40 years now, the system has been encouraging dairy farmers to inseminate on day 60 postpartum, when the cow has a metabolic deficit. This deficit at the time of conception could impact the offspring, especially at the epigenetic level. This project is meant to document the effect of the negative energy balance on the quality of the embryo and to identify ways to improve the fertility of dairy cows.
The beta hydroxybutyrate (BHB) measure was done from blood samples between day 45 and 60 postpartum on eighteen Holstein cows. According to the measure obtained, each cow was classified as low or high in BHB, so as to have at least six cows per group. After an ovarian stimulation process, each cow was inseminated and the embryos were harvested. For each cow, two embryos were transferred in two primiparous cows in order to subsequently determine the persistence of the markers in the biological material. With the EmbryoGENE platform, it was possible to determine the gene expression as well as the methylation status of DNA embryos. The results obtained support the existence of an alteration of the energetic metabolism at the embryonic level, especially by the modification of the signaling pathway of mTOR as well as those of the sirtuins. This alteration appears to result in mitochondrial dysfunction and inhibition of transcription, leading to a reduced activity at a cellular level, probably due to programming of the embryo to use its lipid reserves during severe stress conditions.
iv Table des matières
Résumé ... ii
Abstract ... iii
Table des matières...iv
Liste des tableaux ... vii
Liste des figures ... viii
Liste des abréviations ... xii
Remerciements ... xxii
Avant-propos ... xxiii
Introduction ... 1
Chapitre 1 ... 3
Mise en contexte ... 3
1.1. Évolution de la production laitière ... 4
1.1.1 Bilan sur la fertilité ... 5
1.1.2 La balance énergétique négative en début de lactation ... 11
1.2 Métabolisme post-partum de la vache laitière ... 13
1.2.1 Impacts sur la reproduction ... 16
1.2.2 Effets intergénérationnels de la balance énergétique négative ... 21
1.3 Métabolites indicateurs du statut métabolique ... 25
1.3.1 Les lipides ... 26
1.3.2 Métabolisme des lipides ... 26
1.3.3 Effets des lipides sur l’embryon ... 27
1.3.4 Relation entre le profil métabolique lipidique et la fertilité ... 30
1.4 Le développement embryonnaire jusqu’au stade blastocyste ... 33
1.4.1 Évènements morphologiques ... 33
1.4.2 Métabolisme de l’embryon ... 36
1.4.3 Activation du génome embryonnaire et indépendance génique ... 37
1.4.4 Sensibilité et adaptation de l’embryon à son micro-environnement ... 40
1.5 Programmation embryonnaire ... 41
v
1.5.2 Effets intergénérationnels ... 44
1.6 Modifications épigénétiques ... 47
1.6.1 Compaction de l’ADN ... 48
1.6.2 Méthylation et déméthylation de l’ADN ... 49
1.6.3. Modifications post-traductionnelles des histones ... 57
1.6.4 ARNs non codants ... 63
1.7 Objectifs et hypothèses de recherche ... 65
Chapitre 2 ... 67
Embryonic response to high beta hydroxybutyrate (BHB) levels in postpartum dairy cows ... 67
2.1 Résumé ... 69
2.2 Abstract ... 70
2.3 Introduction ... 71
2.4 Material and methods ... 72
2.4.1 Chemicals ... 72
2.4.2 Animals, management and experimental design ... 72
2.4.3 Measurement of -hydroxybutyrate ... 73
2.4.4 Measurement of non-esterified fatty acids (NEFA) ... 74
2.4.5 Estrus synchronization, embryo collection and implantation ... 74
2.4.6 Preparation of embryos ... 75
2.4.7 Nucleic acid extraction and RNA amplification ... 76
2.4.8 Sample labelling and microarray hybridization ... 76
2.4.9 Microarray data analysis ... 77
2.4.10 Complementary DNA preparation and quantitative real-time polymerase chain reaction... 78
2.4.11 Statistical analysis of quantitative RT-PCR results ... 79
2.4.12 Fragmentation of genomic DNA ... 79
2.4.13 Adapter ligation and DNA digestion by methyl-sensitive enzymes ... 80
2.4.14 Genomic DNA amplification ... 80
2.4.15 Labelling and hybridization ... 81
2.4.16 Array data analysis ... 82
2.5 Results ... 83
2.5.1 Transcriptomic analysis ... 83
2.5.2 Epigenetic analysis ... 89
2.6 Discussion ... 97
vi
2.6.2 Microarray analysis — affected genes ... 97
2.6.3 Microarray analysis — upstream regulator analysis... 98
2.6.4 Canonical pathway analysis ... 99
2.6.5 Epigenetic signature ... 103
2.7 Conclusions ... 104
2.8. Acknowledgments ... 105
2.9 Supplementary data ... 106
Conclusion générale et perspectives ... 125
vii Liste des tableaux
Tableau 1.1 Raisons principales de la réforme d’un bovin laitier et distribution de leurs pourcentages au Canada entre les
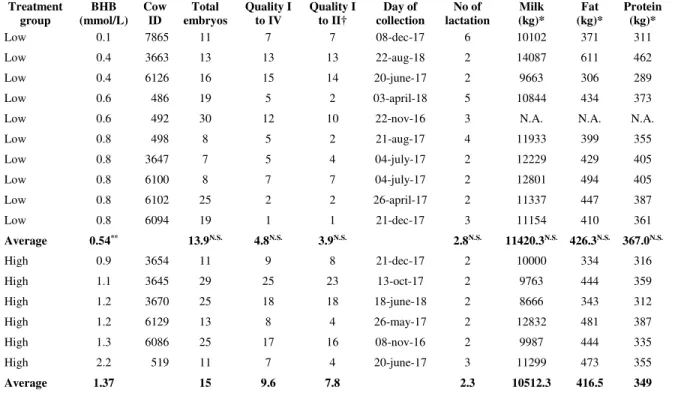
années 2013 et 2016. ... 5 Table 2.1 Number and quality of embryos and milk production for each
cow. ... 873 Table 2.2 Upstream regulators with the largest z scores obtained from
Ingenuity Pathway Analysis ... 87 Table S2.1 Function, relative change in expression (mRNA) and associated
p value from microarray analysis of selected genes. ... 106 Table S2.2 Information on primers used for RT-qPCR validation. ... 107 Table S2.3 List of upstream regulators identified in Ingenuity Pathway
Analysis and their associated activation state. ... 106 Table S2.4 Biological functions of the genes identified in the transcriptome
analysis, suggested by Network Analyst. ... 107 Table S2.5 Identification of the up-regulated nodes using Network Analyst. ... 106 Table S2.6 Identification of the down-regulated nodes using Network
Analyst. ... 107 Table S2.7 Distribution of the DMRs as a percentage of the total EDMA
probes associated with each of the 29 autosomes and the X
chromosome. ... 106 Table S2.8 Biological functions of the genes identified in the epigenetic
analysis, suggested by Network Analyst. ... 107
viii Liste des figures
Figure 1.1 Évolution de la production (kg de lait/vache) des vaches laitières
au Québec de 1975 à 2015. ... 4 Figure 1.2 Moyenne de production en kilogrammes de lait (RHA ; rolling
herd average, de son nom anglais), l’intervalle entre deux vêlages et le nombre de saillies par conception pour 143
troupeaux sélectionnés entre 1970 et 1999. ... 6 Figure 1.3 Relation entre la production laitière et la durée de l’œstrus. ... 9 Figure 1.4 Évolution du taux de conception à la première insémination en
fonction du stade de lactation (JEL). ... 11 Figure 1.5 Représentation des changements associés aux besoins
énergétiques, à l’apport énergétique pour la croissance ainsi que la mobilisation des réserves corporelles de la vache au cours de
la lactation. ... 12 Figure 1.6 Résumé des causes en lien avec la chute de fertilité retrouvée
chez les vaches hautes productrices. ... 16 Figure 1.7 Mécanismes proposés comme agissant sur la qualité
embryonnaire lors de l’ajout de lipides dans l’alimentation des
vaches hautes productrices. ... 29 Figure 1.8 Représentation schématique des étapes du développement
embryonnaire chez le bovin. ... 35 Figure 1.9 Évolution des niveaux d’ARNm pendant le développement
embryonnaire. ... 39 Figure 1.10 Présentation des deux vagues de déméthylation de l’ADN
retrouvées chez les mammifères, associées à leurs périodes respectives du développement embryonnaire. La courbe bleue
est associée au gamète paternel, et le rouge au gamète maternel. ... 43 Figure 1.11 Mécanismes potentiels d’une programmation
intergénérationnelle via la lignée maternelle (rose) ou paternelle
(bleu). ... 46 Figure 1.12 Action des enzymes de la famille des méthyltransférases sur
l’ADN selon ce qu’il s’agisse de la méthylation de novo (a) ou de la réplication de l’ADN (b). Lorsque l’ADN entame son processus de réplication semi-conservative, l’enzyme Dnmt1 ajoute les nouveaux groupements méthyles (rouge) sur le brin
ix
néo-synthétisé (bleu) en répliquant le patron présent sur le brin
original (vert). ... 51 Figure 1.13 Localisation cellulaire de Dnmt1 pendant la maturation et le
développement embryonnaire chez la souris. ... 54 Figure 1.14 Ensemble des voies possibles de la déméthylation active de
l’ADN, ainsi que leurs différents intervenants. La 5mC peut être modifiée chimiquement au niveau de deux sites : soit au niveau du groupement amine (voie de signalisation verte) ou méthyle (voie de signalisation rouge). La 5hmC peut ensuite être oxydée (voie de signalisation rouge) ou désaminée (voie de signalisation verte). L’ensemble de ces produits (Thy, 5hmU, 5fC et 5caC) sont enfin clivés et remplacés par une cytosine (voie de
signalisation bleue). ... 56 Figure 1.15 Différentes modifications possibles de la queue N-terminale du
domaine H3 ainsi que leurs impacts sur l’expression génique. ... 57 Figure 1.16 Collaboration entre les différentes modifications épigénétiques
et leur influence sur la transcription et l’expression génique. L’ajout de groupements méthyles (bleu) sur les sites CpG par les Dnmts permet la répression de la transcription, tout en favorisant le recrutement d’acteurs supplémentaires impliqués dans l’ajout de marques au niveau des queues d’histones. Au contraire, la transcription est permise par l’action de Tet (mauve) qui supprime l’état de méthylation de l’ADN, ainsi que par l’incapacité des Dnmts à se lier aux sites CpG non méthylés. Ce phénomène étant rendu possible grâce aux régions riches
en H3K4me3 retrouvées sur les queues des histones. ... 58 Figure 1.17 Présentation des trois catégories moléculaires permettant la
régulation de l’acétylation des queues aminoterminales des
histones. ... 61 Figure 1.18 Régulation de la transcription par les histones acétyltransférases
(HATs) et les histones déacétylases (HDACs). ... 63 Figure 1.19 Représentation des différentes fonctions jouées par les longs
ARNs non codants au sein de la cellule. Les longs ARNs non codants peuvent recruter différents complexes modificateurs de la chromatine à certains loci spécifiques afin d’en altérer son activité catalytique (a). Ces ARNs peuvent aussi jouer un rôle dans la régulation du processus de transcription par le biais de différents mécanismes en effectuant le recrutement d’autres protéines (b), en jouant le rôle de co-activateur (c) ou via la formation de complexes (d). Enfin, un long ARN antisens non
x
codant peut influencer certains sites d’épissages et entraîner la
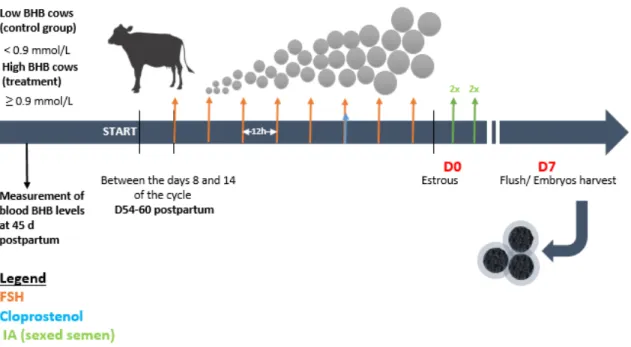
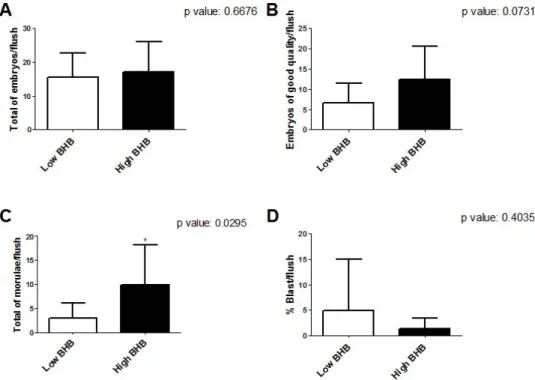
rétention d’introns spécifiques (e). ... 64 Figure 2.1 Timeline of the experimental procedures. ... 75 Figure 2.2 Reproductive performance of cows with low (white) and
increased (black) blood β-hydroxybutyrate (BHB) concentrations. The graph indicates total embryos per flush (A), the proportion of good quality (B), total morulae per flush (C) and the percentage of blastocysts (D). An asterisk indicates significant difference (p < 0.05). Error bars represent standard
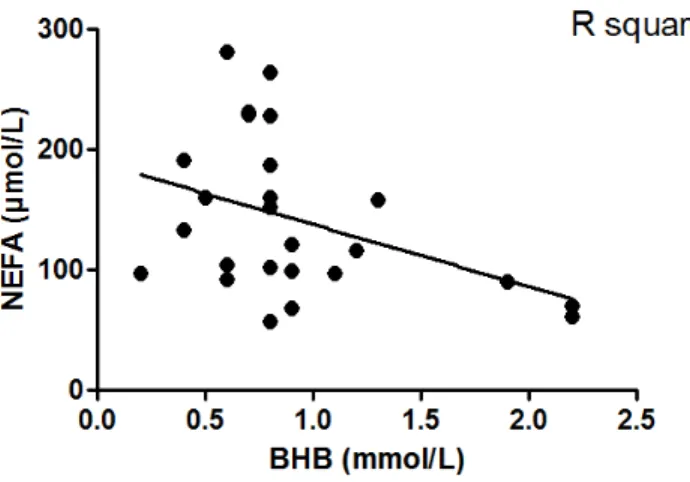
deviations (SD).. ... 84 Figure 2.3 Linear regression of systemic β-hydroxybutyrate (BHB) and
non-esterified fatty acids (NEFA) in the blood of Holstein cows
on day 45 postpartum.. ... 85 Figure 2.4 Systemic non-esterified fatty acids (NEFA, in blood) on day 45
postpartum in Holstein cows with low levels of β-hydroxybutyrate (BHB, white box, n = 16) and in cows with increased levels of BHB (black box, n = 9). Asterisks indicates significant difference (P < 0.01). Error bars represent standard
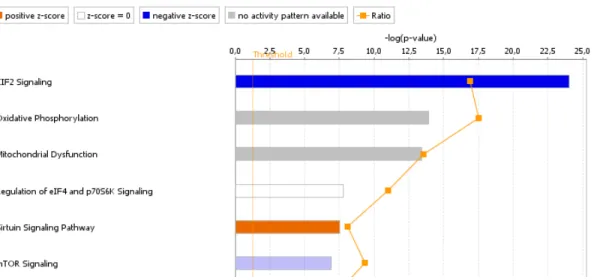
deviations (SD)... ... 85 Figure 2.5 Presentation of the six most important canonical pathways in
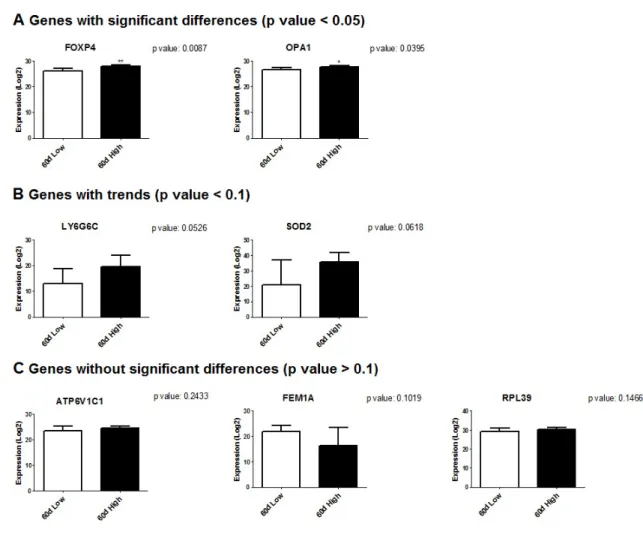
Ingenuity Pathway Analysis from transcriptomic analysis.. ... 86 Figure 2.6 Expression profiles (log2) of seven genes validated by
quantitative reverse transcription polymerase chain reaction (qRT-PCR) in morulae obtained from Holstein cows on day 60 postpartum. Genes are grouped based on their significance (A), trend (B) or absence of significance (C). White: low blood β-hydroxybutyrate group (BHB, n = 6), black: high BHB (n = 6). Asterisks indicate significant difference (*p < 0.05; **p < 0.01).
Error bars represent standard deviation (SD)... ... 89 Figure 2.7 Microarray probes indicating methylation above the background
level in embryos recovered from Holstein cows in the low or
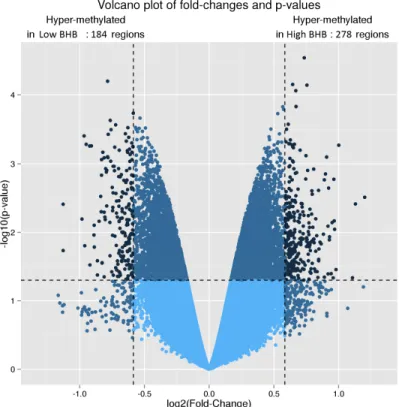
high blood β-hydroxybutyrate (BHB) condition... ... 90 Figure 2.8 Volcano plot of changes in DNA methylation (and statistical
significance thereof, p value) of the Holstein embryo genome, based on microarray results. Embryos exposed in vivo to β-hydroxybutyrate (BHB) at high levels (≥ 0.9 mmol/L) were compared to those exposed to a low level of BHB (< 0.9 mmol/L). Dark dots indicate significant hypermethylation in the control group (upper left) and in the high BHB group (upper
xi
right). The dashed lines represent 1.5-fold changes and the p =
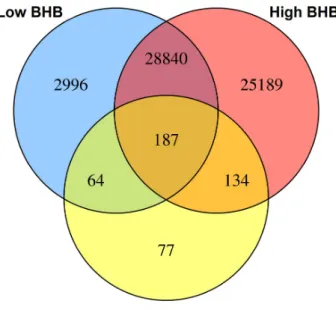
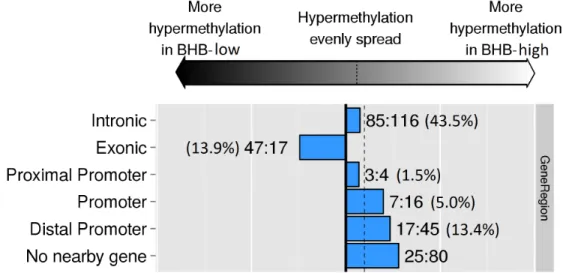
0.05 threshold.... ... 91 Figure 2.9 Differentially methylated gene regions of Holstein embryos
exposed in vivo to β-hydroxybutyrate (BHB) at high levels (≥
0.9 mmol/L) or low levels (< 0.9 mmol/L).... ... 92 Figure 2.10 Circular plot of the combined analysis of the transcriptome
(expression of gene mRNA) and the epigenome (genomic region methylation) of embryos recovered from Holstein cows
experiencing an energy deficit... ... 94 Figure 2.11 Distribution of methylation on chromosomes of Holstein
embryos.... ... 95 Figure 2.12 Presentation of the six most important canonical pathways in
Ingenuity Pathway Analysis from epigenetic analysis.... ... 96 Figure 2.13 Summary of the effects of stress on regulation of components of
the mTOR signaling pathway, an important regulator of cell fate and coordinator of cell growth with energy and nutrient
availability... ... 102 Figure S2.1 Differentially methylated CpG regions of the genome of
Holstein embryos exposed in vivo to β-hydroxybutyrate (BHB)
at high levels (≥ 0.9 mmol/L) or low levels (< 0.9 mmol/L)... ... 120 Figure S2.2 Differentially methylated repetitive element classes of the
genome of Holstein embryos exposed in vivo to β-hydroxybutyrate (BHB) at high levels (≥ 0.9 mmol/L) or low
xii Liste des abréviations
% Pourcentage
4E-BP12 Eukaryotic initiation factor 4E-binding protein
5’UTR 5’ Untranslated Transcribed Region
5caC 5-carboxylcytosine 5fC 5-formylcytosine 5hmC 5-hydroxyméthylcytosine 5hmU 5-hydroxymétryl-uracile 5mC 5-methylcytosine 5mU 5-methyluracil Ac Groupement acétyle
ACTB Actin beta
ADN Acide désoxyribonucléique
ADP Adénosine diphosphate
AGNE Acides gras non estérifiés
AGV Acide gras volatil
AID/APOBEC Activation-induced cytidine deaminase/apolipoprotein B mRNA-editing
enzyme complex
AMH Hormone anti-müllerienne
AMPK AMP-activated protein kinase
ARN Acide ribonucléique
aRNA Antisense RNA
ARNm Acide ribonucléique messager
xiii
ATAD1 ATPase Family, AAA Domain Containing 1
ATP Adénosine triphosphate
ATP6V1C1 ATPase H+ transporting V1 subunit C1
Bax BCL2 Associated X Protein
Bcl-2 B-cell lymphoma 2
BCS Body condition score
BEN Balance énergétique négative
BER Base excision repair
BHB Bêta-hydroxybutyrate
BSA Bovine serum albumin
BWS Syndrome de Beckwith-Wiedemann C Cytosine C16 :0 Acide palmitique C18 : 2, trans-10 cis-12 Acide linoléique C18 :0 Acide stéarique C18 :1 Acide oléique C5 5e carbone CBP CREB-binding protein
CCIL Centre canadien d’information laitière
CCS Compte de cellules somatiques
CCT8 Chaperonin Containing TCP1 Subunit 8
CD437 6‐[3‐(1‐adamantyl)‐ 4‐hydroxyphenyl]‐ 2‐naphthalene carboxylic acid
cDNA Complementary DNA
xiv
CGP Cellules germinales primordiales
CGP Cellules germinales primordiales
COX2 Cyclo-oxygenase-2
CpG Cytosine phosphate guanine
CPT-I Carnitine O-palmitoyltransférase I
CRSAD Centre de recherche en sciences animales de Deschambault
Ct Threshold cycle
DEGs Differentially expressed genes
DES Diéthylstilbestrol
DHI Dairy herd improvement
DLM Domaine de liaison CpG-méthyles
DMRs Differentially methylated regions
DNMT DNA methyltransferase
DNMT1 DNA methyltransferase 1
DNMT3A DNA methyltransferase 3A
DNMT3B DNA methyltransferase 3B
DNMT3L DNA methyltransferase 3L
DR5 Death receptor 5
EDMA EmbryoGENE DNA Methylation Array
EGA Embryonic genome activation
eIF2 Eukaryotic translation initiation factor 2
EMBV3 The EmbryoGENE bovine micro array slide
ER Endoplasmic reticulum
xv
FADH2 Flavine adénine dinucléotide réduite
FEM1A Fem-1 homolog A
FOXO Forkhead Box O
FOXP4 Forkhead box P4
FSH Hormone folliculo-stimulante
GAPDH Glyceraldehyde-3-phosphate dehydrogenase
gDNA Genomic DNA
GEO Gene Expression Omnibus
GH Growth hormone
GH-IGF GH-insuline-IGF-I-glucose
GHR Growth hormone receptor
H1 Histone 1
H2A Histone 2A
H2B Histone 2B
H2O2 Peroxyde d’hydrogène
H3 Histone 3
H3K27me3 Trimétylation de la lysine 27 située sur l’histone H3
H3K36 Histone H3 lysine 36
H3K4 Histone H3 lysine 4
H3K9 Histone H3 lysine 9
H3k9me2 Dimétylation de la lysine 9 située sur l’histone H3
H4 Histone 4
HAT Histone acetyltransferase
xvi
HDAC Histone deacetylase
HDAC1 Histone deacetylase 1
HDAC2 Histone deacetylase 2
HDAC3 Histone deacetylase 3
HDAC4 Histone deacetylase 4
HDAC5 Histone deacetylase 5
HDAC6 Histone deacetylase 6
HDAC7 Histone deacetylase 7
HDAC8 Histone deacetylase 8
HDAC9 Histone deacetylase 9
HDAC10 Histone deacetylase 10
HDAC11 Histone deacetylase 11
HK2 Hexokinase 2
HMGCS 3-hydroxy-3-méthylglutaryl-coenzyme A synthase
ICM Inner cell mass
IGF Insulin-like growth factor
IGF ALS Insulin-like growth factor acid labile subunit
IGF2 Insulin-like growth factor 2
IGFBP-3 Insulin-like growth factor-binding protein 3
IGFBP-5 Insulin-like growth factor-binding protein 5
IGF-I Insulin-like growth factor-1
IP3R Inositol triphosphate receptor
IPA Ingenuity Pathway Analysis
xvii
JEL Jours en lait
KDM5A Lysine-specific demethylase 5A
Kg Kilogramme
KMT Lysine (K)-specific methyltransferases
LDHA Lactate dehydrogenase A
LH Hormone luténéisante
LINEs Long-interspersed repetitive elements
LM-PCR Ligation-mediated polymerase chain reaction
LOC514842 Replication factor C subunit 1-like
LTRs Long terminal repeats
LY6G6C Lymphocyte antigen 6 family member G6C
MAMs Mitochondria-associated membranes
Me Groupement méthyle
MET Transition materno-embryonnaire
MSRE Methyl-sensitive restriction endonucleases
mTOR Mechanistic target of rapamycin
mTORC1 Mechanistic target of rapamycin complex 1
mTORC2 Mechanistic target of rapamycin complex 2
MYC MYC Proto-Oncogene Protein
MYCN MYCN Proto-Oncogene Protein
N.S. Non-significant
NAD Nicotinamide adénine dinucléotide
NAD+ Nicotinamide adénine dinucléotide oxydée
xviii
NAV1 Neuron Navigator 1
NCBI National Center for Biotechnology Information
NEB Negative energy balance
NEFA Non-esterified fatty acids
NTRs Novel transcribed regions
O2- Anion superoxyde
O2* Oxygène singulet
O3 Ozone
OPA One-Phor-All
OPA1 OPA1, mitochondrial dynamin like GTPase
p21 Cyclin-dependent kinase inhibitor 1
p53 Tumor protein 53
PAF Platelet-activating factor
PACS2 Phosphofurin acidic cluster sorting protein 2
pARNi Petits ARNs interférents
PBS Phosphate buffered saline
pCAF p300/CBP-associated factor
PcG Polycomb
PGC1α PPARγ co-activator 1α
PGC7/Stella/Dppa3 Developmental Pluripotency Associated 3
PGF2α Prostaglandine F2α
PHD Plant homeodomain
PI3K Phosphoinositide 3-kinase
xix
PPI Protein—protein interactions
PRC2 Polycomb Repressive Complex 2
PTDSS1 Phosphatidylserine Synthase 1
PUFA Polyunsaturated fatty acid
RAPTOR Regulatory-associated protein of mTOR
Refseq_rna Reference RNA sequences
RepA Regulatory protein RepA
RHA Rolling herd average
RICTOR RPTOR Independent Companion of mTOR Complex 2
ROS Reactive oxygen species
RPD3 Reduced potassium dependency 3
RPL39 Ribosomal protein L39
RT-qPCR Quantitative reverse transcription polymerase chain reaction
S6K1 Ribosomal protein S6 kinase beta-1
S6K2 Ribosomal protein S6 kinase beta-2
SAM S-adénosylméthionine
SF Saturated fat
SGK1 Serum- and glucocorticoid-induced protein kinase 1
SINEs Short-interspersed repetitive elements
SIRT1 Sirtuin 1
SMUG1 Single-Strand-Selective Monofunctional Uracil-DNA Glycosylase 1
SOD2 Superoxyde Dismutase 2
SOD2 Superoxide dismutase 2
ST1926 Adarotene,
xx
STAT5b Signal transducer and activator of transcription 5B
SUV39H1 Suppressor of Variegation 3-9 Homolog 1
TAG Triglycéride
TDG Thymine-DNA glycosylase
TE Trophectoderme
TET Ten-eleven translocation
Tet3 Ten-eleven translocation 3
TF Transcription factor
TSC1 Tuberous sclerosis complex 1
TSC2 Tuberous sclerosis complex 2
UDG Uracil DNA glycosylase
VRK1 VRK Serine/Threonine Kinase 1
Xist X-inactive specific transcript
Zeb2 Zinc finger E-box-binding homeobox 2
xxi
« La connaissance s’acquiert par l’expérience, tout le reste n’est que de l’information. » - EINSTEIN
xxii Remerciements
Je tiens d’abord à remercier mon directeur de recherche, Marc-André Sirard, qui a su me faire confiance et sans qui tout cela n’aurait pas été possible. Merci d’avoir été là, de m’avoir épaulé et d’avoir su mettre ton chapeau de psychologue aux moments propices. Tu as vu en moi quelque chose que moi-même je n’aurais pas su voir, et pour ça, je t’en serai éternellement reconnaissante. Je tiens également à remercier Isabelle Dufort pour sa patience inébranlable. Elle a su supporter mes erreurs, mes nombreuses questions, mes appels de détresse les soirs ou weekends… Je ne pourrais pas oublier nos fous rires, tes commentaires sur mon drôle d’accent et l’amitié que tu auras su me donner.
Merci à toute l’équipe de Marc-André Sirard ainsi que Claude Robert pour leur présence, nos fous rires, nos bières partagées jusqu’à souvent très tard. Votre soutien a beaucoup compté pour moi et a rendu l’environnement du cycle gradué presque tolérable. Un merci particulier à ma collègue Clémence Picard, pour son amitié, son enthousiasme et pour m’avoir ramené Hélène la porcelaine ! Puisses-tu rester encore très longtemps dans ma vie. J’aimerais pouvoir tous vous remercier, mais hélas, je manque de place pour le faire.
Merci à tous les outsiders qui ont su me botter les fesses : Carol-Ann Plourde, ma plus grande amie ; Leslie Desrosiers, ma chère amie et colocataire, qui a souvent dû me réveiller d’une profonde période de procrastination ; ma famille proche qui, même à la fin, ne semblait pas trop comprendre parfaitement ce que je faisais, mais qui m’épaulait avec tout leur amour ! Sans votre aide, j’aurais probablement davantage procrastiné !
Enfin, je tiens à remercier tous ceux qui ont cru en moi et en ce que j’étais capable de faire. Surtout mon père, pour qui je garde une place privilégiée dans mon cœur. J’y suis arrivée !
xxiii Avant-propos
Ce mémoire contient un article scientifique rédigé en anglais dont l’auteure principale est Catherine Chaput. Cette dernière a effectué l’ensemble des manipulations en laboratoire, à l’exception de la mesure des niveaux d’acides gras non estérifiés dans le plasma des vaches ainsi que la décongélation des embryons qui ont respectivement été effectuées par Micheline Gingras et Isabelle Laflamme, techniciennes en travaux d’enseignement et de recherche au département de sciences animales de l’Université Laval. La collecte des embryons à la ferme de recherche de Deschambault a été effectuée par l’équipe du Dr Louis Picard du Service
vétérinaire E.T.E inc. Le matériel biologique a été récolté à la ferme sous la supervision de Janie Lévesque. Les résultats ont été analysés dans leur intégralité par l’étudiante, qui a également rédigé l’ensemble du présent manuscrit. Celui-ci a été révisé par Marc-André Sirard, le superviseur de recherche et coauteur de l’article scientifique ainsi que par Isabelle Dufort, qui était à ce moment, professionnelle de recherche dans le laboratoire de génomique animale de l’Université Laval. Isabelle Dufort a assisté l’étudiante pendant toute la durée expérimentale du projet.
Au moment du dépôt du mémoire, l’article « Embryonic response to high beta-hydroxybutyrate (BHB) levels in postpartum dairy cows » a été accepté avec modifications mineures et sera publié dans le journal Domestic Animal Endocrinology.
1 Introduction
La reproduction est le pivot central de toute entreprise laitière puisqu’elle est la première étape de la production de lait par l’animal. Son importance est donc capitale pour le succès économique et la pérennité de l’entreprise agricole. Or, depuis les cinq dernières décennies, et ce, malgré son importance, la fertilité des bovins laitiers à travers le monde a décliné. Ce phénomène se traduit entre autres par une augmentation de l’intervalle vêlage/première ovulation, une réduction de la durée de l’œstrus, une diminution du taux de succès à la première insémination et une augmentation du nombre de saillies par vache (Butler, 1998; Lucy, 2001; Lopez-Gatius, 2003; Pryce, 2004; Dillon, 2006; Macdonald et al., 2008; Walsh et al., 2011).
Bien que la littérature présente cette dégradation de la fertilité comme étant un problème multifactoriel et complexe, le facteur généralement mis au banc des accusés est la hausse spectaculaire de la production laitière favorisée par la sélection génétique intensive. Au Québec, les vaches modernes sont 30 % plus grosses et produisent deux fois plus de lait qu’à l’époque (Valacta, 2016). Cette attention portée sur les caractères de production au fil des ans a donc permis de passer d’une production laitière moyenne par vache d’environ 7 000 kg au début des années 2000, à un peu plus de 9 000 kg en 2016 (ISQ, 2015). La demande accrue en début de lactation afin de répondre à cet objectif de production est telle qu’il devient difficile pour l’éleveur de remettre la vache gestante, ce qui est notamment expliqué par la mobilisation excessive des réserves de l’animal. Or, ce déficit au moment de l’insémination pourrait alors entraîner des répercussions épigénétiques chez la future progéniture en altérant les marqueurs au niveau du génome pendant le développement embryonnaire. Des phénotypes spécifiques comme la baisse de la production laitière (Gonzalez-Recio et al., 2012) et une diminution de la réserve ovarienne (Evans et al., 2012) ont d’ailleurs être observés chez l’animal adulte ayant subi une reprogrammation embryonnaire.
Le présent mémoire sera donc divisé en deux sections ; tout d’abord, le chapitre 1 consistera en une revue de littérature qui, sous la forme d’un portrait se voulant le plus complet possible, regroupera les connaissances recueillies à ce jour en lien avec le
2
métabolisme post-partum de la vache laitière, la transcriptomique au point de vue embryonnaire ainsi que l’épigénétique. L’accent sera d’abord porté sur la définition de la balance énergétique négative en début de lactation ainsi que son impact sur le métabolisme et les performances de reproduction de l’animal. Une section sera ensuite réservée afin de présenter les différents processus se manifestant lors du développement embryonnaire. Ensuite, les mécanismes impliqués au niveau de la reprogrammation épigénétique de l’embryon seront abordés. Enfin, il apparaît pertinent de présenter une revue des mécanismes épigénétiques pouvant altérer la structure de la chromatine et ultimement la transcription génique. Le chapitre 2 présentera, sous la forme d’un article scientifique, les conclusions tirées en réponse aux analyses transcriptomiques et épigénétiques effectuées sur des embryons provenant de vaches exprimant des niveaux sanguins différents de bêta-hydroxybutyrate (BHB) en début de lactation.
3 Chapitre 1
4 1.1. Évolution de la production laitière
L’industrie laitière a subi une évolution importante au cours des dernières décennies. Afin de se conformer à la demande du XXIe siècle, les fermes se sont modernisées et ont
augmenté leur cheptel. La combinaison de plusieurs facteurs, comme l’amélioration générale de la régie, l’optimisation des rations servies aux animaux et la sélection génétique intensive ont permis l’accroissement spectaculaire de la production laitière, doublant celle-ci en 40 ans (Figure 1.1) (Lucy, 2001).
Tirée, avec permission, de Valacta (2016) Figure 1.1 Évolution de la production (kg de lait/vache) des vaches laitières au
Québec de 1975 à 2015.
La sélection génétique a été particulièrement efficace quant à l’amélioration de la production laitière des bovins laitiers, notamment des sujets de race Holstein, qui est la race principalement retrouvée au sein des cheptels canadiens (CCIL, 2017). Entre les années 1985 et 2003, le gain phénotypique associé à ce critère a été de 193 kg/vache/année pour les États-Unis, 131 kg/vache/année pour les Pays-Bas, 35 kg/vache/année pour la Nouvelle-Zélande et enfin, 46 kg/vache/année pour l’Irlande (Dillon, 2006). Bien que ces pays aient des systèmes de production ainsi que des enjeux climatiques différents, les résultats montrent une diminution générale des performances de reproduction au cours de cette même période.
5
En effet, pour le système de production sous pâturages, exploité entre autres en Nouvelle-Zélande ainsi qu’en Irlande, les études montrent un taux de réussite à la première saillie de 39 à 52 % (Dillon, 2006; Macdonald et al., 2008). Ce taux diminue pour atteindre 30 à 40 % dans le cas des systèmes plus intensifs retrouvés au Royaume-Uni ainsi qu’aux États-Unis (Pryce, 2004; Norman et al., 2009). Falconer (1975) propose deux explications quant à l’existence d’une corrélation inverse entre la génétique et les performances de reproduction. Premièrement, la présence d’un gène pléiotropique agissant de manière antagoniste sur les deux caractères et deuxièmement, l’existence d’un lien entre certains gènes affectant des traits différents.
Enfin, bien que la cause exacte de la chute de la fertilité observée ces dernières années ne soit pas encore tout à fait démystifiée, il est clair que les problèmes de reproduction occupent la première place parmi les raisons d’élimination des vaches au Québec (CCIL, 2017) (tableau 1.1). Avec plus de 42 000 vaches éliminées l’année dernière, les performances de reproduction du cheptel québécois réduisent les exploits économiques de l’industrie laitière qui vaut 17.7 milliards de dollars (CCIL, 2017), tout en augmentant sérieusement les frais d’exploitation.
Tableau 1.1 Raisons principales de la réforme d’un bovin laitier et distribution de leurs pourcentages au Canada entre les années 2013 et 2016.
Adapté du CCIL (2017)
VACHES 2013 2014 2015 2016
Nombre de vaches 698 686 696 737 694 265 691 743
Raisons d’élimination pour la réforme Nombre de vaches Total (%) Nombre de vaches Total (%) Nombre de vaches Total (%) Nombre de vaches Total (%) Problèmes de reproduction 49 815 17.3 % 47 689 18.0 % 44 016 17.7 % 42 251 17.2 % Mammite 35 695 12.4 % 30 569 11.5 % 29 151 11.7 % 27 708 11.3 % Problèmes : pieds et membres 22 399 7.8 % 19 666 7.4 % 18 833 7.6 % 17 643 7.2 % Faible production de lait 20 668 7.2 % 19 159 7.2 % 18 870 7.6 % 17 650 7.2 % Maladie 14 156 4.9 % 13 171 5.0 % 12 677 5.1 % 12 140 4.9 %
6 1.1.1 Bilan sur la fertilité
Tout d’abord, la question la plus légitime que nous pouvons nous poser est la suivante : est-ce que la perception de cette chute de fertilité est bel et bien réelle ? Si celle-ci est réelle, il faut se demander si la production de lait accrue cause irrémédiablement une diminution de la fertilité. Ce qui amène une autre question, à savoir si les capacités nécessaires à une production plus élevée accroissent les exigences sur le plan du métabolisme et de la gestion, sachant que ces exigences ne peuvent alors toujours pas être satisfaites.
L’observation de la diminution des performances de reproduction ne date pas d’hier. En 1929, un auteur du Minnesota dénotait déjà une baisse de la fertilité avec l’augmentation des caractères de production (Hansen, 2000). Plus tard, plusieurs publications documentaient le sujet. Par exemple, Butler (1998) observait une chute de 65 % à 40 % du taux de succès à la première saillie entre les années 1951 et 1996, phénomène qui a été confirmé par plusieurs autres auteurs (Casida, 1961; Dransfield et al., 1998; Lucy, 2001). Ensuite, des données recueillies par Lucy (2001) de 1970 à 2000, à partir de 143 troupeaux d’un système américain d’enregistrement DHI, montrent que la production laitière moyenne des troupeaux étudiés est passée d’environ 6 500 kg de lait à près de 9 000 kg de lait par lactation. Pendant cette même période, le nombre de saillies par conception est passé d’environ 1.75 à 3 et l’intervalle entre les vêlages d’un peu moins de 13.5 mois à 14.8 mois. La Figure 1.2 montre que cette chute des critères de fertilité semble cependant s’intensifier à partir des années 80.
7
Tirée, avec permission, de Lucy (2001) Figure 1.2 Moyenne de production en kilogrammes de lait (RHA ; rolling herd
average, de son nom anglais), l’intervalle entre deux vêlages et le nombre de saillies par conception pour 143 troupeaux sélectionnés entre 1970 et 1999.
Plusieurs auteurs partagent l’idée que la chute des performances reproductives observée au sein des troupeaux laitiers est un problème multifactoriel et complexe (Butler, 1998; Veerkamp et al., 2003). Les interactions entre les nombreux facteurs soulignés rendent difficile la détermination de la raison exacte du déclin de la fertilité au sein des troupeaux (Walsh et al., 2011), bien qu’il semble associer à une augmentation des performances laitières. En effet, il est de plus en plus évident que la fertilité diminue avec l’augmentation de la production laitière, bien que les mécanismes en cause ne soient pas encore suffisamment décrits (Pryce, 2004). Les recherches ont cependant mené à l’identification de causes dites clés qui affecteraient négativement les performances reproductives des bovins laitiers, particulièrement des vaches hautes productrices, qui semblent être les plus touchées par ce phénomène (Walsh et al., 2011). En effet, il semblerait que l’intensification de la sélection génétique sur des caractères de production résulterait en une perte plus marquée de la condition de chair, une dépense énergétique plus importante, l’apparition de désordres métaboliques, associée avec un changement des métabolites sanguins ainsi que
8
le profil hormonal de l’animal. Ces facteurs auraient ainsi un impact négatif sur la capacité de reproduction de l’animal (Taylor et al., 2004; Tanaka et al., 2016).
L’observation de l’évolution de la cote de chair suivant la parturition est une des méthodes permettant d’évaluer l’ampleur d’un déficit énergétique chez l’animal. Butler (2003b) montre qu’une chute trop drastique de l’état de chair (variation basée sur un score d’une échelle de 1 à 5) semble repousser le moment de la première ovulation. Ce retard entraîne alors l’expression d’une plus faible fertilité au moment de l’insémination. De plus, il semble exister un lien important entre le taux de conception et l’ampleur de la perte de condition de chair. En effet, une étude du même auteur indique qu’une perte de plus d’un point d’état de chair, pendant les 5 premières semaines suivant la parturition, peut entraîner jusqu’à une diminution de presque 50 % du taux de conception (Butler et Smith, 1989). Par ailleurs, une étude réalisée par Lopez et al. (2004) montre qu’une production laitière élevée (≥ 39.5 kg/jour) est associée à une diminution de la durée de l’œstrus de 6.2 ± 0.5 h pour les individus au-dessus de la moyenne et 10.9 ± 0.7 h pour les autres (Figure 1.3). Des analyses séparées entre les primipares et les multipares montrent les mêmes tendances et permettent d’isoler l’effet de la parité sur la productivité. Les mêmes auteurs soulignent aussi un plus grand volume de follicules préovulatoires chez les plus hautes productrices malgré une réduction des concentrations sanguines d’estradiol. Cette diminution de la concentration d’estradiol a, entre autres, été proposée comme étant la cause de la réduction de la durée de l’œstrus. Ce phénomène pourrait aussi expliquer l’augmentation du diamètre folliculaire en influençant la sécrétion pulsatile de l’hormone lutéinisante (LH) nécessaire au développement final des follicules, à la production d’estradiol et, subséquemment, à l’ovulation (Wiltbank et al., 2006). De plus, il a été démontré que ces vaches présentent un volume de tissu lutéal plus important, malgré une plus faible concentration de progestérone (Lopez et al., 2005). L’explication la plus probable de ce paradoxe réside dans l’accélération du métabolisme des hormones stéroïdiennes causée par l’augmentation de la consommation de l’animal ainsi que du flux sanguin au niveau du foie (Wiltbank et al., 2006). Ce phénomène, causé par la demande accrue de lait, entraîne alors une chute des concentrations sanguines d’œstradiol et de progestérone, deux hormones impliquées dans
9
la régulation des différents évènements de la reproduction. En effet, lorsque l’animal ingère de la nourriture, le flux sanguin au niveau du foie augmente, tout comme le métabolisme de l’œstrogène et de la progestérone, et ce, selon la production laitière (Sangsritavong et al., 2002).
Lorsqu’une vache est en période de lactation, son foie est environ deux fois plus irrigué et son élimination des hormones stéroïdiennes est 2.3 fois plus rapide que lorsqu’elle ne produit pas de lait (Sangsritavong et al., 2002). De sorte que les changements subis par le métabolisme, suite à l’augmentation de sa consommation, permettent une élimination plus rapide comparativement à la normale. Cet effet est d’autant plus marqué chez les fortes productrices, qui ont des besoins plus importants en énergie et qui doivent alors consommer davantage, influençant l’activité de leur foie à la hausse. L’élimination de l’œstradiol et de la progestérone par le foie varie selon le degré de son irrigation. Ainsi l’augmentation du flux sanguin, causée par l’augmentation de la consommation, entraîne une chute des concentrations sanguines des hormones stéroïdiennes, et ce, même au milieu du pic de production de ces hormones par le corpus luteum (Wiltbank et al., 2006).
Tirée, avec permission, de Lopez et al. (2004) Figure 1.3 Relation entre la production laitière et la durée de l’œstrus.
10
Bien qu’il semble clair que les nullipares présentent de meilleurs taux de conception par rapport aux vaches en lactation (Xu et Burton, 1999; Royal et al., 2000; Lucy, 2001; Peters et Pursley, 2002; Washburn et al., 2002; Gumen et al., 2003; Lopez-Gatius, 2003), le lien entre la production laitière ainsi que le taux de conception est toujours controversé (Wiltbank et al., 2006). Il est en effet impossible de définir un lien clair et précis entre ces deux caractères. En effet, Wathes et al. (2007c) ont effectué des analyses sur un ensemble de données provenant de 500 vaches afin d’explorer la relation entre les caractéristiques métaboliques avant et après la parturition, et la fertilité. Les résultats montrent que les vaches multipares qui ont pris plus de 150 jours à concevoir produisaient en moyenne 13 kg/jour de plus pendant leur pic de lactation que celles qui avaient conçu en moins de 80 jours. Bien que l’ajout de la variable du pic de lactation lors de l’analyse des résultats fût utile afin de prédire la fertilité, l’influence la plus importante sur les performances de reproduction était obtenue lors de la deuxième semaine post-partum. Or, le pic de la production n’était pas atteint avant 5 à 7 semaines. Cette influence sous-entend que ce sont les changements métaboliques associés à la mobilisation des tissus adipeux pour la production laitière qui causent la diminution de la fertilité, plutôt que la production laitière totale de la lactation. Chez les primipares, le pic affecte grandement l’intervalle vêlage-ovulation, mais n’est pas en lien avec l’intervalle vêlage-conception (Wathes et al., 2007c). Un autre facteur à prendre en considération est le moment de la première insémination suite à la parturition (Figure 1.4). Il est en effet logique de penser que la fertilité s’améliore avec l’allongement de l’intervalle entre la parturition et l’insémination. Ce principe repose notamment sur l’attribution d’une période de repos physiologique plus importante. En règle générale, le taux de conception de la vache augmente suite au vêlage, atteignant un plateau à 45 % à 100 jours en lait (JEL) (Bouchard et Du Tremblay, 2003).
11
Tirée, avec permission, de Bouchard et Du Tremblay (2003) Figure 1.4 Évolution du taux de conception à la première insémination en fonction
du stade de lactation (JEL).
En somme, la fertilité chez les vaches laitières semble être le reflet de l’influence cumulative des composants métaboliques, endocriniens et pathologiques post-partum qui sont altérés par la génétique, bien que les liens ne semblent pas indéniables selon les études effectuées. La balance énergétique semble toutefois le facteur ayant le plus grand impact sur la chute de la fertilité, mais les interactions entre l’ensemble des facteurs doivent être considérées et contrôlées (Butler, 2003a). Une meilleure compréhension de la physiologie générale de l’animal parait nécessaire afin de comprendre les problèmes d’ovulation ainsi que de fertilité rencontrés au sein des troupeaux laitiers.
1.1.2 La balance énergétique négative en début de lactation
La balance énergétique est calculée en soustrayant l’ensemble des besoins énergétiques de l’animal (production, entretien, gestation, etc.) aux entrées énergétiques. Cette dernière partie de l’équation représente le résultat de la multiplication de la concentration en énergie par la quantité consommée (Brisson, 2003). Le déficit énergétique peut alors se définir simplement par un excès des dépenses par rapport aux entrées, obligeant ainsi l’animal à puiser dans ses réserves. Selon Brisson (2003), trois conditions pourraient mener à un déficit énergétique, soit un niveau de dépenses élevées, comme une production importante
12
ou un pourcentage de gras élevé dans le lait, un niveau d’entrées faible attribuable à un problème de consommation, comme une faible appétence de l’aliment ou un manque d’eau, et un niveau d’entrées faible attribuable à un mauvais balancement de la ration. Ce dernier critère regroupe notamment une texture inadéquate des aliments, une rumination insuffisante ou une ration faible en énergie. Comme le sujet étudié au sein de ce document s’apparente principalement au premier critère, soit un niveau de dépenses élevé, une attention particulière y sera portée.
La parturition résulte en un changement abrupt au niveau des besoins associés au métabolisme, notamment dû à la demande importante en production laitière. Ainsi, en début de lactation, la plupart des vaches ne peuvent pas combler la totalité de leurs besoins énergétiques par l’apport d’énergie exogène, ce qui se traduit par une altération du métabolisme de l’animal et provoque alors une balance énergétique négative (BEN) (Figure 1.5).
Adaptée de Hoffman et al. (2000) Figure 1.5 Représentation des changements associés aux besoins énergétiques, à
l’apport énergétique pour la croissance ainsi que la mobilisation des réserves corporelles de la vache au cours de la lactation.
13
La BEN débute quelques jours après la parturition et atteint son pic environ deux semaines plus tard (Butler et Smith, 1989; Bell, 1995). Cette période est caractérisée par des changements aux points de vue endocrinien, métabolique et physiologique (Walsh et al., 2011).
Plusieurs facteurs peuvent influencer l’importance et la durée du déficit énergétique en début de lactation. Ces facteurs sont la génétique de l’animal, la cote de chair avant le vêlage, la production laitière, la consommation et l’alimentation (Garnsworthy et Topps, 1982; Bonczek et al., 1988; Grummer, 1995). Chez les bovins laitiers, il semble que l’intensification de la sélection génétique exacerbe ce phénomène (Veerkamp, 1998; Pryce et Løvendahl, 1999). Il y a en effet des évidences que les vaches de haut mérite génétique mobiliseraient plus de tissus adipeux que les autres (Pryce et al., 2001), ce qui est expliqué notamment par l’intensification de la différence entre les dépenses associées à la production laitière plus importante, et l’entrée énergétique (Veerkamp, 1998). En effet, la consommation volontaire de matière sèche semble être un facteur particulièrement important sur la régie de la balance énergétique en début de lactation (Villa-Godoy et al., 1988; Staples et al., 1990). Il est en effet bien connu que les vaches en surpoids au moment du vêlage vont avoir une plus grande perte d’appétit et développer une BEN plus prononcée que les vaches ayant une condition plus équilibrée (Garnsworthy et Topps, 1982). Ainsi, les vaches en surpoids auront une plus grande mobilisation de leurs réserves adipeuses et une accumulation plus importante de triglycérides au niveau de leur foie (Rukkwamsuk et al., 1999).
1.2 Métabolisme post-partum de la vache laitière
Des changements majeurs se produisent au moment de la parturition afin de répondre à l’augmentation drastique de la demande de glucose, d’acides aminés et d’acides gras pour la production laitière (Bauman et Currie, 1980). L’un de ces changements est l’augmentation de la concentration d’acide gras dans le sang ainsi que dans le liquide folliculaire, ce qui est dû à une mobilisation des réserves de gras de l’animal par échange actif (Leroy et al., 2008). Les acides gras retrouvés principalement dans le sang chez la vache en début de lactation sont des acides gras non estérifiés (AGNEs), soit l’acide
14
palmitique (C16:0), l’acide stéarique (C18:0) et l’acide oléique (C18:1) (Leroy et al., 2005b). Les AGNEs sont de longues chaînes d’acides gras de plus de 14 carbones qui se distinguent des triglycérides (TAGs), qui eux, sont estérifiés (Thompson et al., 2014). Ceux-ci peuvent être oxydés au niveau du foie dans le but de produire du dioxyde de carbone, ou partiellement oxydés afin de produire des corps cétoniques ou de l’acétate. Ces derniers sont ensuite expédiés au niveau des tissus afin d’être utilisés comme une source énergétique (Wathes et al., 2007c). Les AGNEs peuvent aussi être estérifiés en triglycérides et s’accumuler dans le foie jusqu’à atteindre une concentration maximale dans les jours suivant la parturition (Bauman et Currie, 1980; Grummer, 1995; Wathes et al., 2007c).
Le principal corps cétonique synthétisé par le foie lorsque les réserves corporelles sont mobilisées est le bêta-hydroxybutyrate, ou BHB, et sa concentration dans le sang est un indice de l’oxydation des acides gras (Wathes et al., 2007c; Fauteux, 2017). Il permet alors de déterminer le degré de mobilisation du foie et l’ampleur du déficit énergétique. L’accumulation de ces corps cétoniques dans l’organisme de l’animal cause une pathologie métabolique nommée acétonémie ou cétose. Au moins 50 % des vaches laitières exprimeront une période temporaire de cétose subclinique dans le premier mois de lactation (Wathes et al., 2007c; Imhasly et al., 2014). On dit que la pathologie est subclinique lorsque l’animal n’exprime pas l’ensemble des signes typiques de la maladie, soit la perte d’appétit, la diminution de la production laitière et l’augmentation de la fermeté des fèces (Duffield et al., 1997). Cette stratégie adaptative de la vache permet de maintenir le taux de glucose sanguin, et ce, malgré une demande accrue de ce métabolite en début de lactation (Wathes et al., 2007c).
Il n’est pas rare de constater de fortes variations des concentrations d’urée pendant la période entourant la parturition. Ces variations peuvent être causées par plusieurs facteurs, dont la stimulation du catabolisme des acides aminés en réponse à une diminution de la disponibilité du glucose, ainsi que l’augmentation de protéines dégradables dans le rumen et un déséquilibre dans le ratio énergie : protéine (Wathes et al., 2007c). En effet, si les rations servies en début de lactation sont trop riches en protéines dégradables et
15
relativement faibles en hydrates de carbone, il se produit alors une accumulation excessive d’ammoniaque dans le rumen (Leroy et al., 2008). L’ammoniaque est ensuite absorbée par la paroi ruminale et convertie en urée au niveau du foie. Or, ce phénomène se traduit par une intensification de la balance énergétique négative en début de lactation et une augmentation sanguine des AGNEs, puisque l’organisme doit éliminer les déchets du métabolisme des protéines, soit l’urée et l’ammoniaque.
Par ailleurs, plusieurs voies de signalisation hormonales se voient modifiées pendant cette même période, notamment celle de la GH-insuline-IGF-I-glucose (GH-IGF) (Lucy et al., 2001). L’hormone de croissance (GH, de Growth Hormone et synonyme de somatotropine) est une hormone pituitaire ayant un rôle considérable sur plusieurs aspects de la lactation, de la croissance et de la reproduction (McGuire et al., 1992; Jones et Clemmons, 1995; Spicer et Echternkamp, 1995; Etherton et Bauman, 1998). Plusieurs tissus en sont aussi affectés (Bauman et Currie, 1980). L’un de ses rôles les plus importants en début de lactation est de coordonner la lipolyse et l’augmentation de la concentration d’AGNEs dans le sang (Bauman et Currie, 1980; Lucy et al., 2001). La liaison de cette hormone à son récepteur (GHR) au niveau du foie entraîne la production d’IGF-I (de son nom anglais, insulin-like growth factor-1), aussi appelé somatomédine C, où il est ensuite libéré par la circulation sanguine. La moitié de sa production est effectuée au niveau hépatique alors que l’autre moitié provient des chondrocytes des cartilages de croissance, des fibroblastes et des autres tissus (Gaillard, 2001). L’IGF-1 possède 50 % d’homologie structurelle avec les régions α et β de la pro-insuline, d’où sa dénomination insuline-like growth factor (Gaillard, 2001). Ce métabolite effectue ensuite un rétrocontrôle négatif sur la glande pituitaire afin de contrôler les niveaux de GH dans l’organisme (Lucy et al., 2001). Cependant, lorsque la vache est en déficit énergétique, cet axe est découplé, ce qui est causé par une diminution de la présence du récepteur à la GH et associé à une chute de la concentration d’IGF-I, ainsi qu’une augmentation de la concentration en GH (Lucy et al., 2001). Lorsque ce mécanisme est couplé en début de lactation avec un faible niveau d’insuline dans le sang, l’environnement endocrinien favorise alors l’action directe de la GH sur la lipolyse et la gluconéogenèse (Lucy et al., 2001).
16
La leptine est une autre hormone métabolique intéressante, qui est en relation avec la balance énergétique puisque sa concentration est fortement associée avec l’état de chair de l’animal, et chute en fin de gestation (Wathes et al., 2007a). En effet, avant la parturition, la concentration de leptine est corrélée avec la cote de chair de l’animal. Elle peut alors être un indicateur de la quantité de tissus adipeux disponible pour une mobilisation subséquente dans le but de supporter la lactation (Lucy et al., 2001; Wathes et al., 2007a). De plus, la leptine semblerait influencer la consommation de matière sèche ainsi que l’apparition de résistance à l’insuline au niveau des tissus (Ingvartsen et Boisclair, 2001).
1.2.1 Impacts sur la reproduction
Il est maintenant clair que la vache subit de profonds changements métaboliques en début de lactation et que leurs impacts varient selon l’importance ainsi que la durée du déficit énergétique. Ces impacts peuvent alors se répercuter sur plusieurs facteurs endocriniens suscités pendant la période post-partum (Figure 1.6). Les sections suivantes résumeront les effets répertoriés de la balance énergétique négative sur le follicule en croissance, l’ovocyte, l’embryon ainsi que le milieu utérin.
Tirée, avec permission, de Walsh et al. (2011) Figure 1.6 Résumé des causes en lien avec la chute de fertilité retrouvée chez les
17
1.2.1.1. Impact sur le follicule, l’ovocyte et l’embryon
Les changements métaboliques subis par l’animal suite à la parturition entraînent des effets sur le cycle œstral, diminuant alors le succès de l’insémination subséquente (Wathes et al., 2007c). Le développement d’un follicule préovulatoire chez la vache prend au moins 40 jours du stade secondaire ou début antral, à l’ovulation. Chez les vaches laitières, la fécondation de l’ovule et les premiers stades de développement embryonnaire peuvent se produire aux environs du pic de lactation, qui se produit généralement entre 70 et 100 jours suivant la parturition (Gonzalez-Recio et al., 2012). Lors de l’insémination à 60 jours, les follicules qui doivent ovuler au moment de la conception entament leurs premiers stades de développement alors que la balance énergétique négative est la plus importante (Wathes et al., 2003; Taylor et al., 2004). La littérature présente de vastes évidences quant à l’impact de la BEN sur plusieurs facteurs dont le développement folliculaire, le développement lutéal et ultimement la qualité de l’ovocyte produit. Cet impact se reflèterait plus particulièrement en atténuant la fréquence des pulses de la LH et en limitant la production d’œstrogène par les follicules dominants (Butler et al., 2003; Leroy et al., 2008), ainsi que d’IGF-I (Diskin et al., 2003). Comme la LH ainsi que l’IGF-I œuvrent de façon synergique dans le but de promouvoir le développement folliculaire (Lucy, 2000), la BEN, en compromettant ce phénomène, entraîne une réduction de la concentration sanguine de l’œstradiol. Ainsi la rétroaction positive moins marquée de l’œstradiol et une ovulation retardée entraîne une diminution des performances de reproduction de l’animal.
Après la parturition, une vague de développement folliculaire se produit en 5 à 7 jours en réponse à un taux élevé de l’hormone folliculo-stimulante (FSH) dans le plasma, et ce, sans tenir compte de la sévérité de la BEN (Butler, 2003a). L’initiation de la vague folliculaire et la formation d’un follicule dominant pendant la BEN ne semble pas être une limitation pour la première ovulation. Toutefois, Beam et Butler (1997) notent trois résultats possibles de cette période de déficit énergétique sur le développement folliculaire : 1) ovulation du follicule dominant (16 à 20 jours post-partum) ; 2) l’absence d’ovulation du follicule dominant suivi par un turnover et une nouvelle vague folliculaire ; 3) le follicule dominant n’ovule pas et devient kystique. Dans les deux derniers cas, il y a une prolongation de
18
l’intervalle vêlage/première ovulation de 40 à 50 jours post-partum. L’ovulation du follicule dominant est toutefois dépendante du rétablissement des pulsations de LH. Cette dernière est favorable à la croissance folliculaire et à la production d’œstradiol (Butler, 2001). Or, la BEN représente un état de sous-nutrition chez la vache qui compromet la sécrétion de LH et empêche l’ovulation du follicule dominant (Jolly et al., 1995). La faible disponibilité énergétique pendant cette période supprime non seulement les pulsations de LH, mais réduit aussi la sensibilité des ovaires à la stimulation par celle-ci (Butler, 2001). La fréquence des pulsations de cette hormone lutéinisante était particulièrement faible pendant la première vague de développement folliculaire chez les individus qui ont développé des follicules dominants non ovulés, comparativement à ceux ayant des follicules où le développement s’est poursuivi jusqu’à l’ovulation (Beam et Butler, 1999). Cette observation est concordante avec une précédente étude des mêmes auteurs (Beam et Butler, 1997) où les follicules se développant après le pic de la BEN montraient une croissance plus importante et un plus grand diamètre.
Les hormones ne sont pas les seuls acteurs touchés par un déficit énergétique. En effet, d’autres métabolites voient leurs concentrations plasmatiques diminuer, notamment le glucose et l’insuline (Beam et Butler, 1999; Butler, 2001). Or, l’insuline est connue pour stimuler le développement des follicules bovins in vitro (Spicer et al., 1993) et in vivo (Simpson et al., 1994). Il semble y avoir aussi certaines évidences quant à l’importance de l’insuline sur la réponse des follicules ovariens aux gonadotrophines, notamment en augmentant l’expression des récepteurs à LH (Frajblat, 2000). Le couple insuline-IGF1 cause aussi une action sur la FSH, permettant ainsi une intensification de la croissance folliculaire en stimulant le recrutement de plus petits follicules (Webb et al., 1999; Armstrong et al., 2001).
L’IGF-I est une hormone pléiotropique composée de 70 acides aminés appartenant au groupe des somatomédines, qui est régulée à plusieurs niveaux dans l’organisme. Les niveaux sanguins de cette hormone peptidique sont primordiaux pour le développement folliculaire (Yoshimura et al., 1996; Gutierrez et al., 2000) et sont directement reliés au statut énergétique de l’animal (Beam et Butler, 1999; Webb et al., 1999). Chez la vache, la
19
chute de la synthèse de cette molécule va de pair avec la réduction de l’expression du récepteur à la GH de type 1A au niveau du foie (Kobayashi et al., 1999; Radcliff et al., 2003b, a; Jiang et al., 2005). Chez les rongeurs, de même que plusieurs autres mammifères, l’activation de la voie JAK-STAT (de son nom anglais, janus kinase/signal transducers and activators of transcription) menant à la synthèse de la protéine STAT5b, entraîne la transcription des gènes de l’IGF-I, la sous-unité acido labile d’IGF (IGF ALS de son nom anglais acid labile subunit) et de son transporteur principal, l’IGFBP-3 (Ooi et al., 1997; Woelfle et Rotwein, 2004; Fenwick et al., 2008). En effet, les IGFBP-3 et IGFBP-5 sont d’importants transporteurs de l’IGF-I au sein de l’organisme. En conditions normales, un complexe ternaire formé de l’IGF, de son transporteur et de l’IGF ALS, prévient la dégradation des IGFs et crée des réservoirs de ces protéines afin d’augmenter leur disponibilité. Ce complexe ne franchit pas la barrière capillaire et prolonge la demi-vie de l’IGF-I, qui passe de 10 minutes pour la forme libre à 12 à 15 heures sous la forme complexée (Gaillard, 2001). Ces complexes ternaires permettent la fixation de plus de 90 % des concentrations totales d’IGF libre (Clemmons, 1997).
Pendant la première semaine post-partum, les concentrations d’IGF-I diminuent de manière drastique, ce qui est connu pour influencer l’intervalle à la première ovulation (Beam et Butler, 1999; Wathes et al., 2007b). Or, au début de la période de déficit énergétique, la capacité de production d’œstradiol par les follicules semble dépendre de la disponibilité de l’insuline ainsi que de l’IGF-I dans le sérum. Suite à la parturition, les vaches qui avaient des niveaux d’IGF-I plus élevés de 40 à 50 % pendant les deux premières semaines avaient ovulé, ce qui n’était pas le cas de celles qui présentaient des niveaux plus faibles d’IGF-I pendant cette même période (Beam et Butler, 1997, 1998). Frajblat (2000) indique que la présence de son transporteur, IGFBP 3, semble tout aussi importante, notamment afin de cibler les tissus, et que sa teneur était plus importante dans le plasma des vaches ayant ovulé lors des trois premières semaines post-partum.
Enfin, certains facteurs provoquant une altération de l’environnement métabolique, comme les acides gras libres ou non estérifiés, l’urée, l’ammoniaque, etc., peuvent alors influencer l’ovocyte ou l’embryon en développement (Sinclair et al., 2000; Lane et Gardner, 2003;