poisscns à
Wemindji, Baie
deJaaœ,
et âDlogie de la:amue
dl Grcerù.aI1d &"ùJS~).par
Bernard M:rin
Méooire de maîtrise présenté à la Faculté des éttdes graduées et de la recherche de l'Université M:Gill cxmne exigence partielle
de la Maîtrise en scieooes.
Départe.rœnt des RessaJrœs Renwvelables COllège MacXlonald de l'Université M:t;ill
Montréal, Québec Décembre 1990 J
J
C;-;','".
,.",,' , . .". .. , rcette
êtWe traite de la oamunauté de poissalS de 1.a ngion de Weminiji, sur la cOte est de la Baie de James. L'utilisation saisonnière de 1. 'estuaire de la rivière Maquatua et des eaux côtières adjacentes par les poiSSOl'lS marins et anadraoos a été décrite en 1987-1988. Cette OCI'lmltlauté de poissons l1altre une faible richesse spécifique (11 espèces) et des variabilités spatio-tmpo:relles ilIportantes dans les captures, œ l'été et 1 'hiver. L'été, l'estuaire est caractérisé par deux espèces marines: le d1aboisseau à quatre cornes (ltfoxooeP.'Jalus guadrioornis) et la lœpén.ie élarrée(l1.n!'I?nus
fabricii).ra
cisoos (Coregonus artectii) et les oorêgoues (C. clupeafonnis) de petite taille « 200 nm) y sont aussi très abc:niants penjant cette l1éne période oatparativement aux eaux côtières o(l la majorité d'entre eux sont de taille ÏllIX>rtante (>300 mn).Dans les eaux ~iêres, le dlaboisseau à épines COJrt:es (M.
soom
ius) , le dlaboisseau arctique (M. scmpioides) et la lOOIUe du Groenlan:l (Gadus.mç)
sont
les espèœs caractéristiques. la nmue du Groenlmù est l' espèce daninante dans l'estuaire perdant l' hiver. l e ; différerxEs dans les clistrihttionssent
influencées, pranièrenent, par des facteurs IilYsiques (tarpératureet
salinité) al 65 ,de la vari.arœ dans les prises peut être expliquéeet ,
deuxièœment, par des oon:li.tioos biologiques(habitat, migration, rep::cxluction) , dépe.niament de la saison.
la mrue du Groenlani, a été étu::tiée plus profoniément au oaJrS de
cette période, ceci afin de clêcrire son éœloqie ainsi que ses caractéris-tiques de vie. Perdant l'étâ, SŒl habitat OOIIesporxl à \.Ble ban:ie de
20 oC. lBs DDrUeS
sent
plus actives la Blit que perdant le jem'. FIl hiver, elles Salt al:xnWrt:es dans l'estuaireet
s'alimentent de façon SClltenue.la période de frai s' éteJù d'avril à juin. la croissance est linéaire et ne diffère pas selon le sexe. la maturité sexuelle est atteinte à trois ans et n'affecte pas le taux de croissance. la repz:oduction a lieu chaque
année et les inüvidus ne dépassent pas l'âge de 9 ans. I.ss carac-téristiques de vie de la oorue du Grœnl.arxl sont différentes de celles de
la plupart des autres poi.ssa1s benthiques de l'arctique: cro.issan;:,e rapide, forte fécxJniité, maturité sexuelle en bas âge et mrl.alité
iIrp:rt:ante. ces caractéristiques peuvent être avantageuses dans les eaux côtières
et
estuariennes de la cOte est de la Baie de James 00. les cxnlltions environnementales soot différentes des eaux arctiques.'lbe ooastal fish OCI'IIIIJJlity of Wenirdji, eastem James Bay, was studied in 1987 an:l 1988 to describe seascmal utilization of the ~tua River estuary
am
the adjacent CXlaStal waters by marineam
anac1ratDJs fishes. Fish diversity was IC7t\f (11 sp.)am
experimental gUInet
catches were highly variable lJeb.'een sites in the estuary am ooastal waters, am also seasonally at a given site. J)]r:in;J SUl'II'Rer, the estua:riœ fishes arenumerically daninated by f-tt.1o marine species, the foorhern sculpin (Myoxooedlalus guadrioomis) an:i the slemer eelblenny (I1mpenus
fabricii),
am
also by juvenile«
200 mu) cisoo (Coregonus artedii)am
whitefish (C. clupeafonnis). In coastal waters, three marine species are
abm:Jant: the shorthorn sculpin (M.
scom
ius) , the arctic sculpin (M. soorpioides) am the Greenlarrl cod (Gadus gmç). In contrast with the estuary, large (>300 m) cisoo an:! whitefish are abm:1ant in ooastal waters irxticatin;J extensive JOOVBnel"lts of these species in James BaydurinJ
the sumner. oistrib.ttion patterns are influenoed, first, by Iilysical factors (water terperature
am
salinity) whldl explained 65 % of thevariatiœ
incatdles
am,
sea:ni, bj biolCXJicalfactors
(habitat d:1oice, miqratiœam
reproduction) c:iepeJW.n;J on the season.Greenlarrl cxxi ~ studiErl in the same period
ta
describe the species'general ecologyam
life history dlaracteristics. Dlrirg the sunmer, they prirx::ipally ooaJpied shallC7ti (2-5 m) ooastal waters. 'lhis habitat was àlaracterizErl by a belt of eelgrass (Zestera marina) at depths of 1ta
3 m, saI inities between 20ta
23AJt,
am
t.atperatures of 3 te 20 oC. Daily t:rap catcheswere
highly variable am nm"e fish 1Nere catqlt atestuary, than in cœstal waters. '1hi.s mvement OOJ:zespc:.rŒ with aàleivement of sexual maturity; spawnirxJ oocurs in April ta J\me. Growth
rate
was
linear for malesam
fanales.'kJe
at first maturity was threeam
rD trade-off was oŒerved between growt:h
am
reproductioo. Greenlard oxl spawned every year am maximJm age was nine years. Greenlam cxxl lifehistory variables differ fran those of JOOSt arctic benthic fishes: they show rapid growt:h, high fecun:lity, law age at first maturity
am
high nmtality. '1bese àlaracteristics nay De .inportant adaptations of cxxite
sustain life in Jane; Bay ooastal waters where env:ira1mental c:x:n;lltions are different than in arctic
waters.
~
... .
USIE DES TABImUX •••••••••••••••••••••••••••••••••••••••••••••••••
I.aISIE DES FIGlRES •••••••••• " •••••••••••••••••••••••••••••••••••••••
mE:F'ACE. • • • • • • • • • • • • • • • • • • • • • toi • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • •
~
...
,... .
~(JtJE Iltl IIJRFA.tT DES 'IIIE5ES ••••••••••••••••••••••••••••••••••••
Emironmental influences CIl seasonal distrihltion of
ooastal
am
estuarine fish assemblages at Wemin:lji, eastenl Janes Ba.y •••••••••••••••••••••••••••••••••••••••Int:rcx:illct.ia1.. • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • iii • • • • • • • • Methods
am
Materials .••.••• Resu1"ts ••••••••••••••••••••••••••••••••••••••••••••••••• DisalSSion ••••.••••••••••••••••••••••••••••••••••••••••• Ref~ •••••••••••••••••••••••••••••••••••••••••••••• Ta.bles: •••••••••••••••••••••••••••••••••••••••••••••••••• FigtreS ... . ~ DE I.IAI~ •••••••••••••••••••••••••••••••••••••••••••••SEcr'ION II Seasonal distrib.Ition, al1Jn:Janoe
am
life history traits of Greenlan:i ccxi, Gadus g;mç, at Weminiji,eastem
James Ba.y •••••••••••••••••••••••••••••••••••••••••••••••••••••Intrc:xJlJc't.ion. • • • • • • • • • • . • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • •
!te:t:Ilc::x3s arx:l ~te.:t-ials. . . • . . . Rest1l"ts ••••••••••••••••••••••••••••••••••••••••••••••••• DiSOlSSion •••••••••••••••••••••••••••••••••••••••••••••• Iœ:f~ •••••••••••••••••••••••••••••••••••••••••••••• Ta.bles •••••••••••••••••••••••••••••••••••••••••••••••••• FiglJl:es ••••••••••••••••••••••••••••••••••••••••••••••••• ~ ET CONCLUSION ~ •••••••••••••••••••••••••••••••••••• ~ 1 ..••••••••••••••••••••••••••••••••••••••.•••••••••••••••••• ~ 2 ••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• ~ 3 ••••••••••••••••••••••••••••••••••••••••••••••••••••••••••• PAGE i
ii
iv
vi
xxii
1 2 5 10 13 19 22 30 34 35 36 39 44 51 58 62 70 81 83 86 89',~ l~" . . . :> ~. h
Je voudrais remercier mes superviseurs Dr. Olristiane Ibial et Dr.
Fred Whoriskey pcm: leurs conseils et leurs critiques au CXllt'S de
œtte
ébde. la ba1ne maràle de l'échantillonnage a étéremue
possible grâce à la c:x>llaboratiœet
au travail d'équipe de plusieurs personnes: steveBaker, Mike RidlardsCll, SUrley Ieam,
DcuJ
Hqle, Alisson Hackney, Nadia"J.bertal
et
Nathalie Doré. cette étuie aurait été difficileœnt réalisable sans la participatim de plu.c;ieurs cris du village de WemWji dont Willie Matches et sa famille quiont
grarrlement oontriblê à JD'l i.ntégra.tion dansla oamunauté. De plus, je rerœrcie le Conseil de barde de weminiji pa1r
l'utilisation de leurs infrastructures (maison, roolotte et entrepôt). Je remercie également le personnel de la station de BiOlogie Arctique et de
l'Institut Maurioe-I.anart:agne du ministère des Pêdles et OCéans pour lr.:ur
S\JRX)rt tedlnique. Je remercie enfin nes confrères d' étuie du CollS;Je Macdonald pcm: leur aide au niveau de l'aspect académique de ma maîtrise. L'\D1 d'entre eux, Benoit Vanier, s'est damé la peine de recueillir peur mi
sur
le terrain les données SUWlélœntaires dont j'avais besoin peur oœpléter certaines de IœS analyses statistiques avec plus de précision. De plus, l'aide d'Ame Evelyet
de M:nica Borobia pc:llr la révision des mamscrits a été très awIéciée. Eh dernier lieu, je ne peux passer saJSsilera! le support finarx:ier direct 01 irrlirect de plusieurs organismes, soit le ministère des Pêcbes
et
Océans, le ministère de l'Diucation du<)lébec, le Fa'Ki poor la fonnation de dlerdleurs et l'aide à la recherche (FCAR) ainsi que le
centre
d'êtmes nordiques de l'Université l>t:Gill.i l
Lm.ΠlB '.IMlDDX
sam:Πl PME
Tableau 1. a;pèces de poissals, myemes des PUE (prises par 22 mUté d'effort) et nanbre total d'in:lividus capturés
à l'aide de filets maillants eJqlérimentaux à chaque staticn durant l'été (1987-1988).
Tableau 2. Moyennes et limites des classes de lcn:jUSUI' des cirq 23 espèces de poissoos les plus al:xnJantes.
Tableau 3. Coefficients de oorrélatioo de Kemall entre les PUE 24 des poisga'lS et les ooordamées des deux premiers
axes de l'analyse DIllticü.Iœnsiamelle oon-mêtrique.
Tableau 4. Prqxntiœ des déploiements de filets maillants de 25
chaque station
awartenant
à d'laque grcupe obtenuspu- l'analyse DIlltidimensionnelle.
Tableau 5. IaJ]UeUrs myemes des trois stades de maturité sexuel- 26 le des cisoos et des coréga~ - nâles
et
fesœlles.Tableau 6. COefficients des deux prenières fooctions caroniques 27 discriminantes non stamardisés p:m: la salinité et
la talpérature de l'eau.
Tableau 7. Moyennes des salinités et des tenpêratures des c~ 28
graJpeS établis â l'aide de l'analyse II'l1lticlimension-nelle.
Tableau 8. Résultat de classification des cirq groopes par 1 'ana- 29 lyse discriminante.
~ŒII
Tableau 1. Efforts d' éc:hantillamage des différents ergins de 62 pêàle utilisés en 1987 et 1988.
Tableau 2. Profaneurs, distan:;,es du rivage et périodes d' 63 utilisatioo des ~ penjant l'été 1988.
Tableau 3. CcIIparaÎSCl'l des caractêristiques Iitysiques des sta- 64 tia'lS d' êd1antillonnage.
Tableau 4. a;pèces de poissoos et rx:mbre total d' intividus qui 65
ŒJt été pêàlés à l'aide de filets maillants
Tableau 5. coefficients de oorrélatiœ de Kemall entre les PUE 66 (prises par \D'lité d'effort) de la nmue du Groenlalrl,
la salinité et la tatpérature de l'eau aux cirq sta-tiœs.
Tableau 6. ~ rtJJYE!IlIleS, fréquerx:les dans la pcpùation, taux 67
de nmtalité, fécorxli.t:2s nDYennes et prqxntions du
Jnnbre total d'oeufs libérés par les 1tOIU9S du Groenlarxi âgées de 3 à 7 ans.
Tableau 7. ~ d'ocx::urrerx::e des proies des nmues du 68 Groenlani prises par les filets maillants
expéri-mentaux pen:Jant l'été et l 'hiver.
Tableau 8. caractéristiques de vie de la mrue du Groenlan:l, de 69 la nmue Arctique et de la
nm:ue
Atlantique.iv
~œ l PAGE
Figure 1. Position géograprlque de llaire dlétme et des stations 30
dléc:hantillomage. les di.st.an::es entre les stations
et
llemba.lchure de la rivière sont iniiquées.
Figure 2. Ordination des ruE (prises par unité dleffort) sur les 31
deux premiers axes obt~..nus par le cadrage nultidinen-sionnel non-métrique. les stations ainsi que la contri-l:ution des espèces ru classes de poissons sont Wiquées.
Figure 3. Palrcentages (%) des ruE des espèces ou classes de 32
poissons poor les cirq gra.Jpes obtenus par 11 analyse multidimensionnelle.
Figure 4. Représentation des centroïdes des cirq 9I":~ selon 33
les deux premières fonctions canoniques discriminantes.
La CXlI'ltri.b.rt:ion de la tenp§rature est irxtiquée.
~œII
Figure 1. Position géograpuque de 11 aire dl ét-Jde et des stations 70 dl échantillonnage. les distances entre les stations et
lleJllbœchure de la rivière sont iniiquées.
Figure 2. lttlyennes des salinités et des ~atures des cirq sta- 71 tians pour les étés 1987 et 1988, et mars 1988.
Figure 3. Variations saisonnières des salinités et des tenp§ratures 72
des cirq stations pen:1ant l'été et 11 autanne 1987
Figure 4. JtkJyennes des ruE (prises par wù:té dleffort) de nmues 73
du Groo..nlan:i prises dans les filets maillants expérimen-taux pour les étés 1987 et 1988, et mars 1988.
Figure 5. Nanbre total de IOOrues du Groenlani prises dans chaque 74
t.raR?e
aux stations 3 et 4 poor un effort de pêdle de 12 heures.Figure 6. Friiquenoes de lon:JUet1I' (intervalle de 25 I1I'I\) des IOOJ:Ues 75
du Groenlan:i attrapées par les trois types dl ergin de pêdle utilisés dans œtte étu:le.
Figure 7. <h1rbe de croissance des IOOrueB du Groenland -mâles et 76 femelles- prises dans les filets maillants expéri.tœntaux.
Figure 8. ftt)yennes des facteurs de oorrlition des roorues du 77 Groenlarrl -mâles et femelles.
Figure 9. PoJrœntages des nmues du
Groenlam,
mâles et femelles, 78(intervalle de 25 nm) qui sont matures en octobre 1987 et mars 1988.
Figure 10. Calrbes de fécon:iité de la IOOrue du Groenlam et de la 79 m:n:ue Atlantique.
Figure 11. M:JyenneS des irrliœs gonado-sanatiques des nmues du 80 Groenlard prises dans les filets maillants expériœntaux.
---
---.---_._---vi
la Baie de James fome la partie inférieure de la plus grarde mer
intérieure du nr:n::Ie (Martini 1986) avec une superficie de 52,000
Jan2
et elle est caractérisée par des ocniiticn; arctiques et su1:rarctiques(runbar 1982). CE!peIŒmt, peu d'êtlxles se salt attardées à dêcrire l'écologie de ses plisscn:; marins et c6tiers, <D'lStituant ainsi \Ul vide
inport:ant dans les ocnnaissances de cette régim. Ré:aiiient, certaines
d'entre
ellesœt
déoontré l' i.nport:.aooe des milieux côtiers de la Baie deJames paJr la productioo primaire (Is;Jerr:Jre et simard 1979; Arderson et Roff 1980; Roff et Iegemre 1986) et benthique (Grenon 1982). Tart:efois, les connaissances sur l' ichtyofaune sont limitœs et localisées à proximité des grarDs estuaires (M:Jrin et al. 1980; Iambert et Dodson 1982;
fobrin et Dodson 1986); c'est poorquoi il est difficile de tirer des
oonclusialS sur II inportance des eaux oOtières poor les poissons. De plus, les dével~...nts hydro-électriques des dernières années dans la région art nalifié le rigime hydrique de deux rivières i.nportantes (la Gran:ie et Fastmatn), ce qui affecta les cxn:litioos Iilysioo-drlmiques estuariennes et oOtières (Messier et al. 1986). Encore une fois, dQ aux oonnaissaœes
limitées sur llidltyofaune côtière, les inpacts de ces c:.tlarxjements l~lont pas été évalués. AjaItons que la Baie de James se situe en zone arctique (I)mbar 1982) mais l'
aR?Ort
en eaux daloes y est très iliport:ant (3.17 x 1011 m3 ; Prinsenberg 1986) ce qui cxn:luit à \D'l mélarge de masses d'eaux froides«
8 oC en été) et saumâtres (20-23 ~) (Prinsenberg 1986) présentant donc des cxnlitions ooéanograpuques particulières. lespoissons marins de l'arctique possèdent des caractéristiques de vie bien
sex-uelle ret:azdêe, faible mortalité et faible fêccnli.t.é (M::Al.lister 1977). A
la Baie de James, ces caract:êristiques peuvent prêsent.er des variantes, en rêpcnse aux cxn:U.tiœs particuliêres dejà mentiCl1ll€!es, qui mêritent d'être documentées.
Nos OCrll'li!Ïssances actuelles peuvent se rêslDner ainsi: les eaux oOtiêres
et
estuariermes de la Baie de James sont friquentées par des poissons marins, anadranes et d'eau doooe (Hunter 1968; Lambert et Dodsal 1982; M:rin et Dodsal 1986). la ridlesse spécifiqueest
faible (22 espèces marines) CDIp1Iëltivaœnt: A d'autres mers dans lenone
(M:rinet
Dodson 1986) et \D'l petit nanbre d' espèces reprêsente unepart
iltportante de la bianasse totale. les espèces de poissals marins les plus aJ.:x:mantes sont la lIm'Ue du Groenlard (Gadus ,gmg), le chaboisseau à quatre cornes(1t{oXoc:leà1alus gyadricomis)
et
le d1aboisseau à épines cnlrtes (M. scorpius) ; l' éc:Dlogie de ces espècesest
peu oonnue dans les eaux oOtières. Trois espèces de salnmidés sont présentes en nanbre awriciable dans la Baie de James: le cisoo (Qxegonusartedii),
le coréga.e (C. clupeaformis)et
la truite nnx:hetêe (salvelinus fontinalis)(Morin gt
Al.
1980). la biologie des poissoos anadranesest
mieux connue p:m' les estuaires i.nport:ants (Ken'pet
al. 1989; Lambertet
Dodson1990) mais l'anpleur de l'utilisation des eaux côtières
est
:iIx:amue. lesestuaires de la rivière la GrarI:Je et Eastmain sur la cOte
est
de la Baiede James sont comus poor être utilisés en été ocmne aire d'alimentation par les poissœs anadranes (M:rin
.et
al. 1981; KsIp §t~. 1989). lesvariatioos jcmnalières
et
sai.scnti.ères de facteurslitYSiques,
m::luant la tatpérature de l'eau et la salinité,ont
une influerœ sur les patrons defane-viii tims biologiques CXIlIIIe la reprcxiuctim
et
l' alimentatiœsœt
aussi iDpn:tantes (tambertet
Dodscm 1982).lB pranier objectif de cette êbxie OCIlSiste den: A dêc:rire les patrons
de distribltim des espêoes de poissa1S fr6quentant. les eaux oetiêres près de Wemirdj i sur la o6te
est
de la Baie de James, rêgial jusqu'à maintenant peu êtmiéet
a
dêtenni.ner l' influeJ'D! des facteurs IitYSi.ques cJangl'êtabli.ssenent de ces patrals de distrfrutiat. U!s informations pennettJ:art de mesurer l' anpleur de l' utilisatioo des eaux oOtiêres par l'ichtyofaune. De plus, face aux nooveaux dévelq:pements hydro-électriques de la prodlaine décennie dans la Baie de James, il
est
pr:imrdial d' au;menter nos connaissances.
L' espèce de poissal marin sélectiamêe poor êtutier plus à fcni les caractéristiques de vie
est
la nmue du Groenlard. Ce choixest
basé sur le fait que cette espèceest
atx::mante sur la ~ est de la Baie de James~in an:i Dodsal 1986)
et
qu'elle a été classée cxmne un poisson arctique (M::Allister 1977).cette
espèce, prodle de la m:n:ue Atlantique (Gadus~), possMe une ba1ne adaptabilité aux variaticms inpntant:es de la salinité
et
de la teqJêrature de l'eau (Scottet
Scott 1988). De plus,cette espèce peut tolérer de bassP.s tarpératures dü à la présence d'une qlycqrotéine agissant ocmne un antigel (Van voorhies et al. 1978). Ces
tolêrames peuvent elCpliquer SŒl suooês dans le noni canadien mais
l'êcologie gênêrale de la nmue
est
très peu connue. Une descriptiCX1 plus détaillée des caractéristiques de vie de la nmueest
den:: nêcessaire palréclaircir
cette
réussite dans les eaux arctiqueset
poor oarprerdre son adaptatiœ aux ocmitions ocêarx:Jgraprlques partic::ulières de la Baie deJames.
I.e secord objectif est den:: de documenter l'utilisatim annuelle des eaux cOtières
et
estuariennes de la rêqioo de Weminij i par la m:>rue dux
ANDERS:ti, J.T.
et
J.C. R>FF. 1980. Sesta1 eoology of the surface waters of Ibma'l Bay. can. J. Fish. Aquat. Sei. 37: 2242-2253.octmAR, M.J. 1982. Arctic marine ecœystems. In Rey, L.R.
am
st:cœhouse, B. (Eds). 'Ille Arctic Ocean, MacMillan, I.J.:nial, p.233-261-GRF1œ, J.F. 1982. '!he mac:robenthic fauna of the Eastmain estuary (James
Bay, ()Jêbec),
before
the cliversien. liaturaliste can. 109: 793-802.lŒHP, A., L. ~ et J.J. IXUD{. 1989. A revisiœ of ooregonid
fish distribrt:ien am al:l.1marD! in eas+..em James-Hldsal Bay. Fnv. Biol.
FÏSh. 26: 247-255.
IN1Bml' , Y. et J. IXŒQ{. 1982.
structure
et rele des facteurs Plysiques dans le maintien des oamunautées estuariennes de poissats de la BaieJames. Naturaliste can. (Rev. Feal. Syst.) 109: 815-823.
1990. n'.fluenœ of freshwater migratiœ en the rE!pl:oductive patterns of anac:lraIoJs pqlll.aticns of cisoo, ~IUS artatii,
am
lake~tefish,
ç.
clweaformis.cano
J. FÏSh. lquat. Sei. 47:335-345.ux;mœE, L.
et
Y. SIMARD. 1979. OOêanograprle biologique estivale et Jhyt:q>larctoo dans le Sld-est de la baie d'Hudson. Mar. Biol. 52: 11-22.MrAILISTER, D.E. 1977. Fea10gy of the marine fishes of arctic canada. W. 49-65. In: sectien II. Marine eoo1ogy. ottawa: Proc. Circunpolar Conf.
on Northern Fmlogy.
MARl'INI, I.P. 1986. cœstal features of canadian Inlarrl
saas.
p. 117-139. InMartini,
l.P. (ed.) • canadian Inlard Saas. Elsevier Oceanograply series v. 44. Elsevier, New-York.rm;sIER, O., R.G. IlGAM et D. R>Y. 1986. Rlysical
am
biologiœl mdificatia'lS in respooseto
la Grarde hydroelectric ClCIIplex. p.403-424. In Martini,I.P. (ed.) canadianInlam
5eas. Elsevier QceaoograPlY series v.44. Elsevier, New-York.M:>RIN, R.
et
J.J. ~. 1986. '!he ecology of fishes in James Bay,Hudsal Bay an:l H\dson strait. p.293-340. In Martini,I.P. (ed.)
canadian Inlarxl 5eas. Elsevier OCleanograply Series v.44. Elsevier,
New-York.
KIUN, R., J .J. ~
et
G. 1OlER. 1980. EStuarine fish OCI'lIIID'lities of theeastern
James-H\dson Baycoast.
Env. Biol. Fish. 5:135-141-1981. 'lt'e migraticms of anadratn1s cisex> (Coregollusartedii)
am
lake wtefish (C. clweafonnis) in estuaries of eastem James Bay. can. J. Zool. 59: 1600-1607.~, S.J. 1986. salinity ard tailerature distr:ib.tticm of Hldsal Bay and James Bay. W.163-186. In Martini, l.P. (m.) canadian Inlard Beas. Elsevier QceanJgra}ily series v.44. tisevier, New-YorK.
R:>FF, J.C. et L. œ;m;:m:. 1986. lbysioo-dlanical and bioloqical oceanograPly of HOOson Bay. p. 265-291. In Martini, I.P. (ed.).
canadian Ward Seas. Elsevier
00ean0cJraPlY
Series v. 44. Elsevier, New-YOI'k.SOJl'I', W.B.
et
M.G. SCDrr. 1988. Atlantic fishes of canada.cano
Bull. Fish. Aquat. sei. 219: 731 p.VAN VOCRIIES, W.V., J.A. RAYI«H> et A.L. DE.VRIES. 1978. Glycqlt'oteins as
bioloqical
antifreeze
agents in the cxxi, ~ QEÇ, (Ric:baIàsal).xii
Iê œmidat a le droit, après awrooatim du ~, d' in::lure
0CIlIIIe partie intl!grale de sa thèse le texte d'un cu de plusieurs nanus-crits pœvant être samù.s A un périodique paIr p.lb1icatim. Dans ce cas, la thèse doit toojClll."S se c:alfonner aux autres exigerDeS expliquées dans ce docmnent
et
du matériel acHitiau'lel (clc::lrlnh; brutes, détails sur de l'êquipanentet
plan d' échantil1amage) peut être nécessaire. Dans toos les cas, une int:roductiooet
\me ooo:::lusioo doivent être in=luseset,
quam
il Y a plus d'un manuscrit, \.Dl cœmentaire de liaison, \meintroduc-tioo
et
une 00l'X:1usion CŒI1I.1IleS sont nécessaires. Un sinp1e recueil de manuscritset
des réi:npressiCl1S de papiers p.tb1iês ne sont pas ac:x:ç-tables.Même si l' aàlltiat de manuscrits cn-rédigés par l'auteur
et
d'autres persames n'est pas interdite IXJUI' l'examen, le caniidat est averti d'iniiquer clairement l'aJ;pJrt de dlaam durant le travail; les directeurset
les oo-auteurs devront tâtrJigner de la validité de ces déclarations devant la amité oral. Il faut ooter que dans ce cas la tâd1e de l'ex-aminateurexterne est
d'autant plus difficile.carme
le pennet la Faculté des étmes graduées,cette
tllèse CXl1tientdeux mamsc:rits. la première section, axée sur la oamunauté de poissons, sera samdse A "Ehv.iraJnenta1 Bio1ogy of Fishes" et la seoorde, portant sur la 1IDt\1e du Groenlard, le sera à "canadian Journal of Z00logy". ont
co1labaré à ce projet en
tant
que cn-auteurs C. Hldonet
F. Whoriskey.xiii de la rêdactim. la pertinence de cette êtlœ se fcxœ sur le fait que les
infœ:maticms ClCI'1terIles dans ces deux maJ'IlSC['its salt ŒAlVe1les.
i,
BIvirœ.mal .inf1.uemes
cm
fJPianü distribJti.œ of aastal. anI2 lHBIDCl'lClf
Estuarine
am
ooastal fishes inhabit adynamie, often unstable enviraJDent (Felleyam
Felley 1986). Estuaries in particular, ocx:upy a transiticmal positicn between fresham
salt water envircnnents (Felleyam
Felley 1986; Knox 1986). Fish distrihttiCllam
al::A.Jrdaooe in these habitats are determined firstlyl:7t
IilYsicalam
dlemical factors inclOOirg salinity, tarperature, ~, tide, wavesam
win:l (Ham 1982),am
secxnUy by biological functions such as migration, reproduction, feeclirg
am
habitat selection (Blaberam
Blaber 1980; Darrlo 1984; Whitfield 1988).Estuarine enviromnents are especially inportant as nursery areas for juvenile marine
am
anac1ratws fishes (Clarià;Je et al. 1986; Knox 1986).Juvenile Atlantic sal.nal (sall'lD salar) (Power et al. 1987; amjak et al.
1989)
am
sane ooregadd species (Bordam
Eric::ksa\ 1989; I(errp et al.1989) may also use estuaries as a transitimal marin} envircnnent l::efore or durin) tl1eir migratiCll ta lOOI'e saline waters. 'lbeir estuarine stay may
facilitate their adaptatiœ
ta
marine ocniitioos.Coastal waters
am
estuaries of HOOson Bayam
James Bay are frequented by freshwater, diadratoJsam
marine fishes (Hunter 1968;Iambert
am
Dodsal 1982; ftm'inam
Dodson 1986; fb'inet
~. section II).MJst of CllI' kmwleàJe of fish CXI'IIIIlJ'li.ties frou the:se ragions canes fran stuties evaluatin} the envircnnent:al inpacts of hyàro-elect.ric develcpnent of large rivers. '1he fL~ oammrl.ty of eastem James Bay is characterized
3
by a low diversity of anadr.'aID.1s (lake wtefish, QrecgIlS clupeaformis, cisoo, ~. artedii
am
brook traJt, Salvelinls faltinalis) arr:i marine(Greenl.alù axi, Gadus g;mç
am
MVoxoœdlalus SAlo) species (MJrinam
Dodsal 1986) 0 Migratmy patterns of the anadraIoJs species have beenrelatively ~ll documented CH=rin et al. 1981; !trin am Dodsal 1986) 0
cisoo am lake whitefish were reported
to
migrate am feed in ooastal waters of James Baydurin.J
the sunmer, with juvenile wtefishœinl
ab.nJant in
estuariœ
durirg this period (M:>rin et al. 1981; Kenp et al. 1989). Juvenile cisoo distributiCX1 is rŒ yet documented in James Bay. CCltparatively little is known of the seasonal distribItion of marine fishes in cœstal waters (lt:>rinam
Dodsal 1986) 0 Alse, the :ilrport.aooe of smallerri
vers, estuaries
am
ooastal waterste
marineam
diadranous fishes is poorly knawn.In James Bay, daily am seascl'al variatiœs in Iilysical factors, incllXiirg terrperature
am
salinity, play an inportant role indeterminirkJ
fish distribltiCX1 (Lambertam
Dodsa1 1982; Prinse.nberg 1986). Alorç with these dlaracteristics, biological ft.Jœtioos suàl as reptoductionam
feed.iJ'g are also 1:hc:u;Jhtto
infl\lel'Da the distriblticn of anadralIJus a.-n marine species (I.ambert an:l Dodson 1982; MJrin et al. section II).'!he objective of Ws stuiy was
te
describe the fish assemblages (CXllpœed of imividuals >100 1lITl fork lerçt:h) of the Maquatua River estuaryam
the adjacent shailCM ooastal waters at different times of the year. 'l11e influence of water terrperature arr:i salinity a l distril:utiol1identi.fy if cbaracteristic fish assent>lages occured at given sites.
Based (Xl previoos kmwleëÇe of the biology of fish species
am
their envir:trtnelrt, we t.ested two predictions: 1) that juvenile whitefisham
cisoo are mxe abJrdant in the estua:ry nursery area than in CX>aStal waters am 2) that daily am seasonal variatioos of salinity an:! water t:.enperature were largely influen::il'g fish distribltioo.5
'!he study site
was
located near Wemin:lji (53°00'N, 78°S0'W), a smal1cree
OC'II.1ll.Ulity situated at the 11DIth of the Maquatua River in easternJames Bay (Fig. 1). '!he east coast of James Bay is characterised by
Preœmhrian cryst.alline rocks an:i is dotted with
many
islamsam
reefs (Dionne 1980). '!he Maquatua estuaryam
its contigurus coastal waters are sha1low «15 m). '!he sediIœnts are mainly silt, clayam sam.
Eelgrass (Zestera marina)am
laminaria beds are CCITIIDn in rlle coastal area (Roche 1982).'!he Maquatua river was danIned in 1983, aba.lt 0.5 km upstream fran the junction with James Bay. Mean annual discharge before danmi.n;J was 15.8 m3 s-l (Wemirrlji Bani COUncil, urplblished data). No infonnation is available on the disc:harqe after
dantni.rr:J.
In the sunmer, freshwater inflow fram the Maquatua river ani lani runeff maiIltain highly stratified waters in the inlarx:l part of the estuary. Distally the estuary is weIl mixed by wim an::}tidal currents. Fstuarine waters exhibit varirus canbinations of salinities (0-23
wt)
am
tenperatures (5-23 OC) due te short term c.hargesin wim directions an:i velooities, current patterns, freshwater runeff
am
tidal anplitmes. seasonal variations aIse oc:x::ur. Marine coastal waters of James Bay typically have salinities of 22-23wt
am
water t:enperatures rarely rÎSE' above 15 OC in SUll'IIEI'. '!he ice-free pe.riod inOOIXÜ.tions are mre stable than in S\lI'II.œr because of the presence of a 70
an ice CXNer an::l reduced fresh water i.np.rt:s (lti:>rin B. personal
oJ:servations). Tides in James Bay are semidiurnal with a lOOderate
anpl~tude (0.5 te 1 mi Godin 1974).
Biological
am
Plysical samplinjsanplirg took plaoe between J'une 1987
am
July 1988 at five stations loœted in the Maquatua estuaryam
its adjacent coastal waters (Fig. 1). '!he stations wre positioned alon:j a gradient of Iilysical factors (t.eIrperatuream
salinity) fran the estuary te marine coastal waters. AlI statior., had similar depthsam
slopes of the lx>ttan. Fishes were oollected usinJ experiIrental gill nets (50 m IOIlj; 6 panels of mesh sizes25,39,51,64,77
am
102 mm) deployed periodically (1-2 week intervals) at the 5 stations fran June-<)ctober 1987am
May-July 1988. In winter, \.1Jl:ler-ioe gill nettirg was oorxlucted in early March 1988; however only stations 1,2,3 wre accessible (Fig. 1). Gill nets \'Vere set perpenlicularto
shore at depths of 2 te 3 m, for 24 h periods. Occasionally bad weatherprevented their recovery
am
the set lasted 48 h. In winter, precautions 'vere taken te avoid gill nets freezinj te the umer-ioe surface.Accord-injly, they were set at 3 m depth because of the presence of the 70 cm ice
00Vet.". '!he overall fishing effort was 76 gill net
days
in S\lJ11IœI'am
10gill net days in winter. catparisons of gill net catches fram stations 4
am
5 in October 1987 were net made due te excessive fishirg tiIœs caused7 AlI fish caujlt were identified, cnmt:ed and measured (fork leBJth ±
0.5 lIIIl) an:! weighed (± 0.5 g). cat:dles
are
expressed incatch
per unit effort (œJE), with a Wlit effart œÏD1 24 heurs. Mean 1eD)th of male andfanale Witefish an:! ci.soo were CXIIplted at three maturity stages: (1) irrInature (ga'lads very sanall am threadlike) (2) develq>irr;J (gonads
enIargÏD1 for
spawnin:J
in the current year) am (3) mature (gametes in amature fonn) based al the classificatiCVl of Snyàer (1983).
water tatperature an:l salinity profiles were made with a SID-12 and Bec:kman RS5-3 salinaneter OOIlCXIIÛtant with gill net
sets.
Data analyses
'!he cat:dles of the JOOSt al:llmant (>200 irxtividuals taken) fish species were divided in three len:fth classes: small, medium
am
lazge. Size categories 'Nere determined l7j the 33.3am
66.6 perœntile values of theCUIIIll.ative size distribItion of a11 sarrples poolai for eadl species. 'lhis division
was
arbitrarily dlœen dueta
cm- poor knowledcje of PJPÙationstructure
am age at maturity of many spec:ies ineastern
James Bay. It aIse provided an objective criteriata
divide fish species ~ 3 a:JUa1 size classes. Eac:h size classwas
t.reated as one variable for thestatistical analyses.
follltivariate teàmiques were used
ta
test tmether the gilinet
catches at the five stations were biologically classifiable into distiB::tgrcups obtained
l:7.i
the analysis woold rep:esent spatio-teq;oral entities of fish species or size classes that wa.üd Ile faull tcgether because of s.imi.lar enviraIœntal calStraints, or biologicalinteractialS.
'lbe data analysis procedures were:
A simi1arity matrix of the different species
am
size classes of 9i11 netsets
was oonsb:ucted usirg the Ia.ùczynski coefficient of similarity (IBgemream
Isgemre 1984):S = 1/2 (W/A
+
W/B)W= SUIn of minimal al::l.n'œro! of all variables 'h= SUIn of aIl variables for the firet sarrple B= SUIn of aIl variables for the secxnl sanple
'Ibis semi.-œtriœl ooefficient OalSiders the al:sence of a species or a size class in a qil1
net
set as an abserœ of informatiCl'l rather than as an element of similarity (lRqeRkeam
IBgemre 1984). 'Ibis matrixwas
t.hen subjected
ta
carplete linkaqe clusterirg (5orensen 1948)am
to
a ncnnetric nulti.di.mensional sca1inJ (Kruskal 1964) • (Dp1ete linkage clusterirg was usedto
obtain graJpS ~ manbers of the clusters arehighly related (Le. species or size classes that oooccur together) (I.egenjre
am
Isgen:lre 1984). Nonœtric nultidimensional séalin) was usedto
aCXXllJlt for the ~ of many dooble zeros in the catch data am talepzesent the relatiCl'l between abjects in a spaoe of reduced ctiJne.nsions (I.egenjre
am
I.egenire 1984). 'lbe two methods were oanbined sc that clistin:::t qroops obtained by the derm'ograns of the cluster analysis were9 nCIlDetric scalin;J. 'l1le classificaticn ptooedl1res Wlen CCIIIbined with ordinatiCll teàmiques sunrnarize the b.ùk of data d:Jt:ained
am
identify the iDpartant species in eadl bioloqical association. '!he first axes of the Ol'dinatiCll EDq)lained a high pIcplrtiCJ\ of the variarœ otserved (Iegenjream
Isgerdre 1984). 'Ihus, in arderta
distin;Juish m:D} the dlaracteristic species or size classes in the ordi.natiCl'lam
clusterirg of qill net sets,~ic Kerdall cmzelaticn cxmficients were cx:atp1t:ai between a11 variables
am
the fint axes resultin;J fran the rxrmet:ric scalin;J. Eadl g:rwpwas
then characte.rizEd aCCOI'diB!J ta its fish species or size classes. '!he classificaticn of qillnet
sets obtained by the nultivariate analyseswas
oarpared ta spatial distrihrtioo of the sanplin;J stations am Iilysical àlaracteristicsta
aoc:œnt for thestructure
observed.Tc evaluate the inport:ance of water salinity
am
terperature on the distribltiCl'l patternsoœerved,
a discriminant analysis was performed onthe matrix of bottan ard surface salinities
am
tenperatures mea.sum:i cxn::urrently with the qill net sets. Discriminant fuœtions resultiDj fran the analysis pennitted usto
identify the relative iJrportance of the principal P'lYsical factors in discriminatirg between the grŒJpS obtaiœi by the clusterirgam
ordinaticn analyses.Experimental qill net catdles
A total of 11 species were caught in eJ!PeI'imental gill net catàles (Table 1). cisex> daninated gill
net
catdles durin:j the S1.JIIIDe[' wleGreenlani cod.
was
predaninant durin:j the winter. Atlantic lunpfish (qrclcpt:e.rus IlntplS) am Atlantic he:rrin;J (Clupea hare[gus) were excluded frou the analysis ta avoid the spJrioos effects of rare species « 5% oa:urrence). Five species whose total catdleswere
greater than 200.iniividuals were divided in sma11, medium
am
large size graJpS (Table 2).For the otller species, fish of a11 sizes wre pooled into one variable.
Ordination
am
cluster analysis'!he oanbinatlOll of cœplete linkage clusterirg
am
noranetric nulti-dinensional scalin"J analysis establisœl 5 distin::t grtXJpS dlaracterized by different fish species am/or size classes (Fig. 2). '!he level of similarity between qill net set::. within eadJ. groJp was higher than 0.40.Ordination of the gill net sets
am
the superinposition of the gra:q:s were only represented on the first two axes of the nultidimensional scalirgi no separation of the groups was evident alorg axis 3.'!he first axis was significantly oorrelated with 14 variables
am
theseoom
with 6 (Table 3). '!bus, the first bic axes were Ïltp)rtant for the fonnatioo of the 5 groopsam they
can be characterized by different fish----_._ ... _._-_ ... _._--_ ..
_~._-11
species
am
size classes. '1be highest cxn:xelatia'lS lIIere fa.uxi with fC1Jrhorn saùpin (It{oxooeglaluscmadricomis)
am
cisoo. Of the five species divided in size classes, aùy the caregarl.ds exh.ibitedoorrelatia1S of qlpCSite signs aJID"g size classes, irdicatirg differenoes
in
distribltial with size. Groups Aam
B were foxmedl7:i
gill net sets cx:mi.n;J fran staticms 1 an:i 2, in the estuary of the Maquatua River whereas gralpS C, DamE tNere daninatedbf
gill net sets fran stations 3,4 am 5in
ooastal waters (Table 4; Fig. 2).Small (juvenile) caregarl.ds, especiaUy cisoo, wre allJnjant at
estuarine statiCllS (gra.1pS A
am
B) (Fig. 2,3; Table 5). 'The fourhom sculpin was the daninant species regardless of size in groop B. '!he slen:le.r eelblenny (Ilmpenus fabricii) was also present in significantnumber in grcA.Jp B. '!he shorthorn sculpin (Myoxoœg,alus sooIpius), the
arctic sculpin (M. soorpioicles)
am
the arctic staghom sculpin (Gymnocanthus tricuspis) oc:nlI'ed CIlly in groups C,D am E. catches in group C representedtransitory
c:xnlitiœs betweenestuarine
(A,B)am
ooastal (D) assemblages slooe the nœt ab.udant species (cisco, Greenlmn cxxl) w.re equally represented • Group D oœprised medium an:l large inti viduals of bath ooregonid SJ:'.ECies. Gm.Jp E
was
dlaracterizedb.i
theGreenlarxi cod. '!he slerxier eelblenny an:i the foorhorn so.ùpin were scarce
in
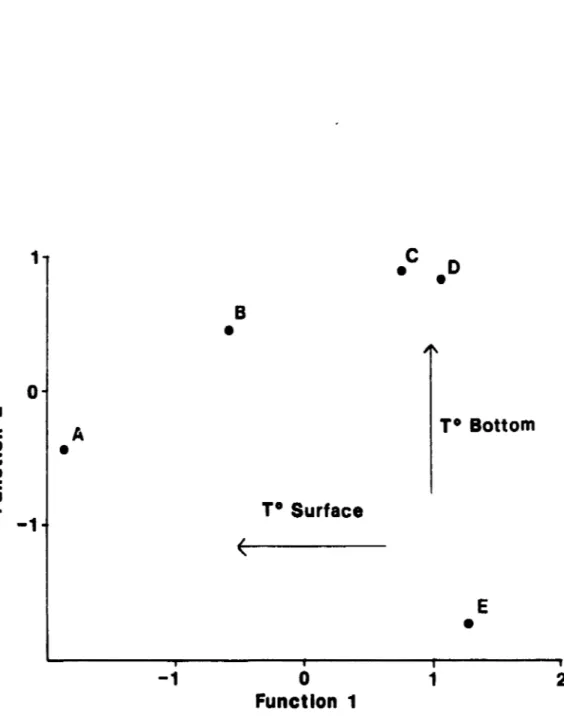
stations 3,4,5.function coefficient (Table 6). 'Ibis function clearly separated gra.Jp; A an:! B fran a11 the ethers (Fig. 4). GraJps A
am
B are formed by gi11 net setsoanirY:J
fran stations 1 anl 2, in the estuary of the Maquatua River. 'lbesestaticrs had low surface salinity
am
high surface teJrperature (Table 7).'lbe seocni discriminant :fUnction elq)lained 40.7 % of the variability, an:l
bottan tenperature had the highest coefficient (Table 6). '1his function primarily separated graJp E fran aIl ether groops. GroJp E was fOl.'11Bl by
qill net sets
oan.in:J
fran a11 stations with low water t:elrperatures. M:>st sets were fran. winter anl early sprirq. overa11, 63.5 % of the grouped cases were correctly classified by the discriminant analysis (Table 8). Classification results of the discriminant analysis were highest for qrrups Aam
Eam
smaller for grcAlp C. Incorrect classltication ofcatd1es of groop C fish as qroop 0 may have oocured due ta similarities in their Ptysical characteristics.
---13
'!he Maquatua estuary exhibits inport.ant spatio-tarporal variability in lilysica1 dlaracteristic:s,
wàl
oon:espads with high variability in fish captures. 'lbe superpositial of gradients in timeam
space was best studied usirç lIIÙtivariate statistiœ,web
provide an objective means to dete1:mine the transitiŒl points within the CDltinuum of Iilysical an:lbioloqical conditions.
'!he 5 groop; obtained by the l'Illltivariate analyses inlicated different spatio-taipJral utilization by fish of inshore waters in the Wemin:lji
area.
'!he spatial cœponent was inpJrtant durin]sunmer
ta differentiate between groops A,B,Cam
D. Fadl groopwas
characterizEd by gil1 net sets fran ale or two statiCXlS. Groops Aam
B, CUip:sed of 9i11 net sets at statioos 1 an:l 2 in the estuary of the Maquatua River were characterized by marine fishes am juveniles of coregonids whidl presmnab1y have a highto1e:ran;:,e for ICM salinities
am
high t.eJtpe.ratures (Mrinet
al. 1981).'!he daninant marine species in the estuary
durm;,
the sumner was thefwrhom saùpin, siJnilar to ether estuaries in James am 1b:isal Bays (foiJriu
am
Dodsal 1986). 'Ibis species tolerates low salinities,am
lives in coastal estuarine waters in arctic an:l sub-arctic regions (Scottam
scott
1988). AlI three size classes of fcmhoom sculpin ~ present in the estuary SUJ]eStin;J that this species is primarily an estuarine resident. 'lbe ether marine species present in the estuary in surrmer, slerder eelb1enny, is also wiclely clistrib.rt:ed in arctic am sub-arctic waters (Scott an:l Scott 1988). '1heir preserre in brackish waters may be anirdicatiœ of the species' flexible habitat requiraDents
am
their high adaptability far estuarine ocnii.tialS.'lbe p:reseroa of :iDnature eisco am whitefish in large ~ in the estuary durin) the SUl'l'IDeI' is an aspect of ooregadd distribltial that bas
been poorly described. It bas been reportai that in sunmer juvenile la1œ wtefish were very abnDant at the river muth
am
in the estuary of thela Granle (Morin et
Al.
1981)am
Little Whale rivers (Ke1tp ~ jÙ. 1989),lrtbile inmature ei.scxl were
scarœ
in the estuaries. 'lbe general biology of tbe two species showsstrorx.J
similaritities. Beth spawn in fresh waters inthe faH (lotJrin
am
Dodson 1986). Larvae hatdl the followin;J sprirgam
may be flushed ta Jaœs Bay1:7.i
the heavysprirr:.J
runeff (Ochman am Dodson 1982). However, eisoo have semi.b.loyantam
pelagie larvaeweb
are nm-e susceptible ta he carried into James Bay than lake wtefish larvae whicht:errl
to
remain in the spawnirg gravelam
use estuaries as arrursery
(M:Jrin
et
al. 1981). '!he transport of coregonid larvaewas
stlxli.ed aùy inlarge estuaries
am
in the oontig'lOOS ooastal waters of eastem James Bay.rur
results shoWed that juvenile eisoo are abJrdant in estuarine waters,in oontrast ta previoos stmies. A possible explanatiœ of this differenoe is that larval eisoo, flushed into offshore waters of James Bay, retum ta
coastal
waters tutnet
necessarily ta the same area franweb
they came. 'Ibis dispersal WI:All.d give juvenile eisco aocess ta inshore habitats suchas the Maquatua estuary whieb does net have a local spawni.rq pcp1lation
due to danI:nirg. 'lhe pcpù.atioo of eisoo associated with the Wernirdji area ooo.ld ranain ta
rear
until ad1ei vin) sexual maturity. 1.bis issuworted
by15 (Ibbl urplblished data). ~ achievin:J first mawrity, they waùd return te their natal river far spilW1'lÏn), alth::u#l E!VideIxle of hanirJj ranai.ns te
ta
delralSuatai (Mlrin §t §le 1981). Because of the hydroelectric damwàl
bloàcs acoessta
spilW1'lÏn)grcuœ,
a naturally reptolucin:j~llatiŒl in the Maquatua River is laàd.n;J. '1he law spawnin;J activity associated with the Maquatua River is also obvious fran the fact that juveni.le lake wtefish are rot abJrœnt \tben CCI'l'pU"ed
ta
ether sites(e.g.
Ka't1?
gtAl.
1989).Adiitialal factors cnùd also oart:rib.rt:e
ta
the high abJrrjance ofjuvenile cisco in estuarine waters. '1he utilizaticn of estuaries as a transitmy an:! nursery habitat by other salloonid species bas been
described (Qmjak et ~. 1989; Power
et
al. 1987). Itwas
SUC}';JeSted that thisPlase
was .i.Jrportantta
alla« grar~ of the fishta
a sizewere
~imal osnDregUla.t:ory abilities are develq8i (Omjak et al. 1989). 1his
hypothesis oould explain larvae
am
juvenile Witefish ab.lrœrD!s in estuarine waters (lot>rinet
al. 1981). Hawever, osrooregulatory abilities waùd have ta develcp early in life for cisoo, si.ra! the larvae are flushed into salt water. '!he retum of juvenile ciscota
inshore watersafter their dispersal
ta
offshore areas of James Bay cannat be t:.ernai a transitmyJi1ase.
Rearin:j in brac1dsh waters may decrease enerqetic oosts associated with osrooregulaticn, pennittin:J the investment of extra ~'" in grcwt:hdurinJ
the first years tmere nmtality is .i.Jrportant (Lambert am DodSŒl 1990). '!he small size of these fishes wrul.d also make themwlnerable ta predation by larger fishes oocurin:j in ooastal waters. JUvenile cisco were foord in stanadls of Greenlan:l axl
am
sculpins(llrin B., urplblishai data). By DDVin;J ta the estuary they might
avcicl
these marine predatars ani take advant.ar)e of the availability of a largefood SlJR)ly (lb'in gt
Al.
sectim II).Fish in groops C am D came mainly fran statims 3,4,5 am shared the same marine species (sborthorn, arctic
am
arctic staghom saùpins, thelatter recmded for the first t.ime in James Bay). Sculpins fOl'1ll an inport:ant part of the fish OCIIIl'ImÎ.ties in eastem Jarœs Bay (!tJrin am Dodsœ 1986). 'lbese marine fisbes rarely venture into the estuMy
(excl\din;J the sborthorn sculpin) presumably because of their pref~
for oold
am
salty waters (scottam
ScxJtt 1988).Group D, formed of fish calJjlt at st" ... ",ticns 4
am
5, was daninated bylarge
am
medium cisco, inoontrast
to SlIall cisc::Dweb
are limitai to the estuary. '!he presence of léU"""~ ciscx>am
lake whitefish at statioos 4am
5, the farthest staticns fran the river nnrt:h, probably Wicates the utilizatim ofcoastal
waters asfeedirYJ
gra1I'rls durirg the sunmer. Ia1œwhitefish were repœ:ted to make extensive mvanents into ooastal waters, b1t no sudl informatim
was
available for adult cisoo (ltrin etAl.
1981).'1hus, jlÙ:Ji.rg fran a.JI' capture of mature cisoo at least 15 km fran the
river lIDlth, bot:h species
were
distribltedam
mved extensively in coastal. waters. similar ooastal mvementswere
observed for arctic cisooin the Beaufort sea (Craig 1984; Bord
am.
Erickson 1989).---
---17 '1his grœp cblJllEl1ts short as we11 as lCDJ tallot.al
àlarJ1es
inan
distribJti.cm. In Slner 1 this species isaœent
franwann
estuarinewaters ~in ~
Al.
sectim II)am
JIK7YeSte
colder coastal waters (statiCllS 3,4,5). In winter they are al::aJrdant in the estuary (statiat 1)am
use this habitat as afeedin:J
and spawnin} grcAllÙ (}i:Jrin et ~.sectiœ II).
water t:alperature was the ncst significant Iilysiœl factor measumi that explained the variability ~ the grcAJpS. '!he separation of groops A, B
am
graJpS C, D, E based CIl surface talperature ildic:ates thatjuvenile cisoo
am
lake whitefish preferwarm
tatperatures. catparable resultswere
obtained for arctic cisoo in the Beaufort sea (Fec:belm ~ al.1983;
craig
1984). '!he infl~ of oottan terrperature in separatirg groopE
fran
aIl ether grc:K.Jp; is stra-gduri.n:J
thesunrner
in ooastalwaters,
ardbec allies minimal in winter Wen biological factors are playirg a JlDI'e
i.Jrportant raIe (lttrin
et
al. sectia'l II) • '!he overall OO:t:1ectclassifiœtioo of 65 % of g:rc::AJpS by the discriminant analysis based on salinity and tatperature, is a gcxxl irdiœtiat of the ~ of these factors
to
explain the distrib.lticn of the fish assEIIblages in the wemi..rnji arm. '!he pcor performa1'D! of the classifiœtioo ofsets
forgraJp C (33 %) in::li.cates a transitioo point in the gradient of Ptysic:al
am
biological OOIlÜtiCl'lS. '1his law pe.rœntaqe might also be due to relatively lcrg net-set duratia'lS (24 h) in CCIlpaI'ison ta possible short tenu variaticm in Iilysical factors (tide, weather) which oooldnet
berecxmBi. '!he influerœ of };ilysiœl factors is maximal in the sunœr W'1en bioloqical factors suàl as repr:oductive migratiCl'l for oaregadds are
neg1igible. In winter, reproductive
am
ovetWi.nter mvements by CXJrega'lidam
marine species accoont far a large anomt of variabi1ity in the gi11 net catches sirœ water tJ:mp!ratures are haIIxjeneals anaJj statiœs (see secticm II).In sunmary, the distr.ibltiŒl patterns of the fish assemblage in the Wemindji area are prirx::ipa11y influeI'XBi l:7j t.arperature
am
sa1inity especially in the SUl'IIIIeI' , with biological factors such as lIlJVeDeIlts of--
---_.
19
BIABœ., S.J.M.
am
T.G. BIABER. 1980. Factors affectirg the distr:ihttion of juvenile estuarine ard inshore fish. J. Fish Biol. 17: 143-162.BC.H), W.A. ard R.N. œIClŒXfi. 1989.
sunmer
stmies of the nearshore fish CX'III1'I1JÛty at RUllipsBay,
Beaufortseacmst,
Yukal.cano
Tech. Rep.Fish. Aquat. sei. 1676: vii +102 W.
Cl.ARIDGE, P.N., I.C. POI'l'ER
am
M.W. HARDISIY. 1986. Seasooal dlarçes in JIDVBDeI1ts, ~, size CXilp:lSitim an:) diversity of the fish faunaof the
severn
estuary. J. Mar. Biol. Assac. U .K. 66: 229-258œAIG, P.C. 1984. Fish use of roastal waters of the Alaskan Beaufort 5ea: A review. Trans. Am. Fish. Soc. 97: 61-63.
aKJAK, R.A., E.M.P. aJAIJI1IΠani M. EEFARS. 1989. Dc:Jwnst.&.""eal1l IOOVemellts an:1 estuarine residerDa
l7:i
Atlantic salmn parr (salnD salar).cano
J. Fish.}.quat.
Sei. 46:1466-1471-DANOO, P.R. 1984. Reproduction in estuarine fish. pp. 155-170. In: Fish reproduction strategies
am
tactics. G.W. Pottsam
R.J. wootton(eds. ). ü:n:lon: Academic Press.
DlOONE, J.C. 1980. An cutline of the eastern James Bay coastal environ-lllel1ts. In S.B. M:=Cann (Ed.), Geel.
surv.
ofcan.,
paper 80-10 : 331-338.FEXlJE[M, R., W. NEILL ard B. GAI..tDlAY. 1983. Te.nperature preference of
juvenile Arctic cisoo (Coregonus autumnalis) fran the Alaskan Beaufort
Sea. Bioloqical Papers of the University of Alaska 21: 24-38.
FEr.U.Y, J.O.
am
S.M. FEl.l.E.{. 1986. Habitat partitiOllirç of fishes in an urban, estuarine bayaI. Estuaries 9: 208-218.GODIN, G. 1974. '!he tides in east-...em am western James Bay. Arctic 27: 104-110.
GREN:fi, J.F., 1982. '1he macrobenthic fauna of the Fastmain estuary (James
Bay, Q.lêbec), before the diversion. Naturaliste cano 109: 793-802. lIUNrER, J.G. 1968. Fishes
am
fisheries. Al. 360-378. In: C.S. Beals(ed.),
scien:Je,
Historyam
HOOsonBay. canada
Dept. El'lergy, Mines &ResaJrœs, ottawa.
JENSEN, Ad. S. 1948. Cl:>ntrib.rtion to the ichthyofauna of Greenlard. Skrift. U. Zool. Mls. Kobenhavn 9: 182 W.
1Œlt1P, A., L. BœWIafEZ dl'rl J.J. IX)[)SŒ. 1989. A revis ion of ooregonid
fish distrihltion
am
ahlrrlanoe in eastem James-H\xlson Bay. Env.m;)X, G.A. 1986. Estuarine ecosyst.ems: A systars çproad1.
œc
prœs.
Boca Ratal, Florida. 289 g>.I<RUSl<AL, J.B. 1964. Na1metric nultidimensional scalin;J: a rrumerical
~. Psychaœtrika 29: 115-129.
IAMBERI', Y.
am
J. J. ~. 1982.structure
et rôle des facteurs fbYsiques dans le maintien des oamunautées estuariennes de poissonsde la Baie Janes. Naturaliste cano (Rev. Ec:x>l. Syst.) 109: 815-823. 1990. Influern! of freshwater migration on the reproductive pattems
of anadrcIto.ls pep.ùations of cisoo, Coregonus artedii,
am
lake~tefish, g. clupeafomis.
cano
J. Fish. 1\qUat. sei. 47: 335-34S. LmnIDRE, L. an:i P. m.;ENDRE. 1984. Ecologie rn.nnérique, 2ièIœ Mition.Tane 2: la structure des données écx:>logiques. Masson, Paris et les Presses de l'université du ~ébec. viii+ 335 g>.
ux;mœE, L. an:i Y. SIMARD. 1979. Ooéarlograprle biologique estivale et (.'hytq>lancton dans le Sld-est de la baie d 'HlxJson. Mar. Biol. 52: 11-22.
MARl'INI. loP. 1986. Olastal features of canadian Inlan:i Saas. g>. 117-139. In: Martini, loP. (ed.) • canadian Inlan:i Saas. Elsevier Ooeanograp1y series v.44. Elsevlar, New-York.
MANN, K.H. 1982. Ecology of coastal waters: a system
awrœch.
Sbxlies in ecx>logy v.8. university of california. Iœ An;Jeles. 250 W.lm)SlER, O., D. ROY
am
R. ~. 1985. Evolution du mercure dans ladlair des poissons. Réseau de surveillaJX)e écologique du carplexe la
~ 1978-1984, Société d'énergie de la Baie James, Direction lrgeniérie et envi.ronnerrent. 170 g>.
K>RIN, R. am J.J. IX)[)OCN. 1986. '!he ecx>logy of fishes in Jaœs Bay,
HI.Xlson Bay
am
Hlxlson strait. g>. 293-340. In:Martini,
l.P. (ed.)canadian Inlan:i Seas. Elsevier OoeanograP'lY Series v. 44. Elsevier, N.Y.
MJRIN, R., J.J. ~
am
G.roolm. 1980. Fstuarine fish <XII1l'llIlities ofthe eastern James-HOOson Bay coast. Env. Biol. Fish. 5: 135-141.
1981. r.rhe migrations of anadrœnls cisoo
(coregonusartedii)
am
lake~tefish (Coregonus clupeafomis) in estuaries of eastem James Bay.
cano
J. Zool. 59: 1600-1607.OCHMAN, S. am J.J. IX)[)OCN,. 1982. eutposition am structure of the larval
am
juvenile fish CXITI1Illli.ty of the Eastlnain Riveram
estuary, James Bay. Naturalistecano
109: 803-813.21 PCMFB, G., M.V. PatlER, R. tufAS
am.
A. ~. 1987. Marine migratiœ of Atlantic salDal fran rivera in Urgava Bay, Queœc. Am. Fish.soc.
syq,.
1: 364-376.PRINSENBEK;, S.J. 1986. 5alinity
am
t.e:lpe.rature clistril:utialS of Hlù;onBay
am.
James Bay. g:».163-186. In: Mlrtini, l.P. (ed.) canadi.an In1arxiBeas. Elsevier Ocearxxj.raply series v. 44. Elsevier, N. Y.
KXHE Ltee, 1982. Etudes ~ de la cOte
est
de la Baie de James. 'ltIDe III. la vêgêtatiœ littorale.RaRxrt
prêsent6a
la SEBJ(Société d'Ehergie de la Baie de James).
sa:œr,
W.B. and M.G. SCDIT. 1988. Atlantic Fisbes of canada.cano
Bull. Fish. ~t. Sei. 219: 731 Al.SNYDER, D.E. 1983. Fish eggs
am
larvae. W.165-197. In: L.A. Nielsen andD.L. Jc:tmson (eds.) Fisheries t:.echni.ques. American Fisheries SOciety.
sa<AL, R.R.
am
F.J. RJiLF. 1981. Biaœt:oxy. 'lbe principlesam
practice of statistics in biological research. 2m edition. W.H. Freeman, SanFrarx:isal. 859 g>.
OORENsm, T. 1948. A method of establi.sh.inJ g:raJpS of equal anplitude in
plant sociology basecl on similarity of species content
am.
its ap-plicatioo ta analysis of the vegetatioo a l Danish UililOllS. Biol. Skr. 5: 1-34.WHITFIEID, A. K. 1988. '!he fish OCII'IlIlI1ity of the swartvlei estuary and
the influence of food avaibility CIl resoorce utilization. Estuaries
Table 1. Means (± SO) of elCpel'imental qill
net
catàl per \DÙt effort (CPUE) foreacn
speciesdurin:J
SUllIDe.I' 1987am
1988. '!he total l'lUI'IiJer (n) of fishfor eadl species
am
the pIqxntia'l of the total catàl (%) are in:ücated.statia'l n % catàl Species 1 2 3 4 5 Marine g. gmç 2.4 4.3 10.4 4.4 7.2 731 23.8 (3.2) (3.8) (5.4) (4.0) (5.6) M. quadrioornis 3.6 9.9 2.8 1.3 0.2 468 15.2 (5.5) (6.5) (2.4) (1.2) (0.4) , M. SOOIpius 0.0 0.6 5.0 2.6 7.4 369 12.0 !
1
(0.0) (0.7) (3.0) (2.5) (4.4),
~ M. scotpioides 0.0 0.0 1.4 1.3 0.5 66 2.1f
(0.0) (0.2) (1.7) (1.5) (0.8)~
J.,. fahricii 0.4 1.3 0.1 0.1 0.0 48 1.6 ~ " (0.7) (1.6) (0.5) (0.2) (0.0)t
g g. tricuspis1
(0.0) 0.0 (0.0) 0.0 (0.9) 0.5 (0.7) 0.4 (0.6) 0.5 33 1.11
~. lUlplS 0.0 0.0 0.0 0.1 0.2 7 0.2 (0.0) (0.2) (0.2) (0.3) (0.4) ~.haremus
0.0 0.0 0.0 0.0 0.0 2 0.1f-i
Anadl:'atols (0.0) (0.2) (0.2) (0.0) (0.0)r.
~. artprlii1
7.4 8.2 12.2 10.8 14.7 1080 35.1 (18.4) (9.9) (8.6) (7.6) (12.0) ~. Ql~ar:mis 1.4 2.7 3.0 2.3 0.3 227 7.4 (2.9) (3.3) (2.1) (1.9) (0.8) ~. fmtinalis 0.2 0.0 1.0 0.6 0.2 46 1.5 (0.4) (0.0) (1.3) (0.8) (0.4)l
23
Table 2. Size classes limits
am
nvaans for the fi ve JOOSt aturnant species. stamarà deviatiCXl is iniicated in braclœts.Size class (DIU)
Species StaIl Medimn large
~. artedii size interval <202 202-269 >269
(n=1057)
x
161 240 292 s (28) (19) (22)Si·
gmg s.i. <259 259-300 >300 (0=691)x
217 287 343 s (35) (11) (46) M. auadricomis s.i. <225 225-240 >240 (n=447)x
200 232 253 s (27) (4) (10) H. soozpius s.i. <221 221-253 >253 (n=293)x
195 237 285 s (22) (04) (10) ~. cl\paformis S' .L <252 252-300 >300 (n=215)x
206 278 333 s (30) (13) (28)Table 3. KelDall oorrelaticn coefficients between
the gill
net
œJE's of fish species or sizes an:lthe ooordinates of the first 2 axes fran the rametric nultidimer.sicml scalin;J. Signifiœnt correlatiCllS are urr".erlined (p<0.05).
Species
g.
artedii Small Medium largeg .
.Q9ilÇ Small Medium LargeH.
quadrioornis Small Medium LargeM.
soorpius Small Medium Large ~. clupeafonnis Small Medium LargeM.
SOOlpioidesy.
trieus,pis ~. fontinalis L. fabricii Axis 1 -0.109-ihm
-0.421 0.561 0.465 0.214 ~ 0.027 -0.173 -0.306 -0.306 -0.140 Axis 2 -0.244 ~ -0.147 -0.154 -0.038 -0.024 0.119 -0.085 0.028 0.024 0.180 0.290 0.212 -0.062 0.181 -0.09325
Table 4. P:r:cportioos of the
sets
fran eac:h statial bel~in;J to eadl group defined fran the 1IIlltivariate analysis. NlDnber of gillnet sets
(n) is irxlicat.ed. Highest prqlOrtions are urderlined.station n 1 2 3 4 5 A 23 ~ 35 0 0 0 B 15 13 87 0 0 0 C 20 5 10 55 30 0 D 19 0 0 0 53 47 E 19 26 10 47 5 11
•
Table 5. Mean leB]th (JlI1l) of cisoo (COt'egOp\ln artedii) and laJœ
wtefish (C. clupeafonnis)
oorrespadinJ
ta
the three maturity stages. standanl deviatiœ is iniicated in braclœts as well as the number (n) of fish measured in each grwp.Male Female Maturity stage 1 2 3 1 2 3
ç.
artediix
197 265 298 189 245 288 s (43) (32) (23) (35) (38) (28)n
108 192 15 140 177 140ç.
clypeafonnis 260 289 330 233 293 337 (65) (37) (46) (46) (30) (41) 21 40 11 21 33 24 26Table 6. tJnst.an3ardized œnoniœl discriminant furx:tion coefficients for salinity
am
tenperature. Percentage(%) of explaÎned variarx::=e is also :i.rdiœted.
8alinity (wt) SUrface Bottan Tenperature (OC) SUrface Bottan % variaJx,e explained 1 0.111 0.121 -0.289 0.129 56.0 F\mct.ion 2 0.061 -0.093 -0.003 0.292 40.7 27
Table 7. Mean (± SO) salinity
am
tenperatuœ recorded for the sets belCDIin:Jta
ea.c::h graJp deteJ:mined by the mltivariate analysis.salinity (ppt) 'l'E!t'perature ( OC)
n surface surface bottan
A 23 8.1 20.4 12.4 7.1 (7.2) (2.2) (4.0) (2.9) B 15 17.5 21.7 13.8 8.7 (3.2) (1.0) (3.4) (2.9) C 20 20.9 21.5 10.7 9.6 (4.8) (1. 7) (4.1) (3.5) 0 19 22.3 22.3 10.6 9.4 (0.6) (0.6) (2.5) (1. 7) E 19 15.3 22.7 3.5 2.3 (9.6) (1.3) (3.9) (2.9)
---_ .... _-_ .. _-_ ...
__
.. _--.-._ ... _ .... _ ... _- ... _ .. _----_ ...._--_._---
...Table 8. Classification results of the discriminant analysis. F\1rcentaqes (t) of abjects that were classifiecl in the actual groop. 'lhe nuruber 00 the diagonal (umerlined) repIesent the pzcporticn of
sets
oorrectly classified.Actual groop n A A 23 70 B 15 13 C 21 0 D 19 0 E 18 0
Predicted groop membership
B 13 60 10 5
o
c
13 7 21o
o
4 13 48 68 11 Eo
7 10 5 29,
,
/ / , HUDSON BAY ': .'..
~
p ~:~ ~ ~ 1 2 3- 4 , 1 1 1 km JAMES BAY..
AQUATUA RIVER ~., .... , .:.: .... :;~.~. DISTANCE FROM RIVER MOUTH STATION fkm) 1 2 3 45
3 8 11 14 16 w oFig. 1. Gec:graJ;hica1 location of study area ani position of saupli.Jg
staticns. Distarv::&; of the stations fran the river IOOUth are also given.
Axis Il
1.5
10.5
0-0.5
-1 • • • -1 - ---.~---,G.
trlcuspls
M.scorploldes
&ML M.
scorplus
ML
G.oglc
JlL C.Irtedii
D•
• • ••
•..
• • • ••
• •-0.5
o
L. '.brlcll
SM M.quadricornis
• C . •rt.dU
••
8t.2 ••
• ••
•
• • • ••
0.5
1
) 31 SML C.artedll
L C.
clupelformls
SMG.
•1.5
ogac 2 Axis1
Fig. 2. Ordinatioo of the gill net CPUE's 00 the first two axes obtained fran the noranetric DlÙtidimensiooal scal.in;J. GraJps
CA
to
E) fran the oc:atplete linkage clusterin;J are overlayed. statioos with the highestprcp:ntian of



![Table 5. Mean leB]th (JlI1l) of cisoo (COt'egOp\ln artedii) and laJœ wtefish (C](https://thumb-eu.123doks.com/thumbv2/123doknet/7661581.238604/45.924.159.753.508.821/table-mean-jli-cisoo-cot-artedii-lajœ-wtefish.webp)