Optimisation, étude de la cinétique et dimensionnement de la production des biopesticides à base de souches de Bacillus thuringiensis isolées du sol Libanais
Texte intégral
Figure

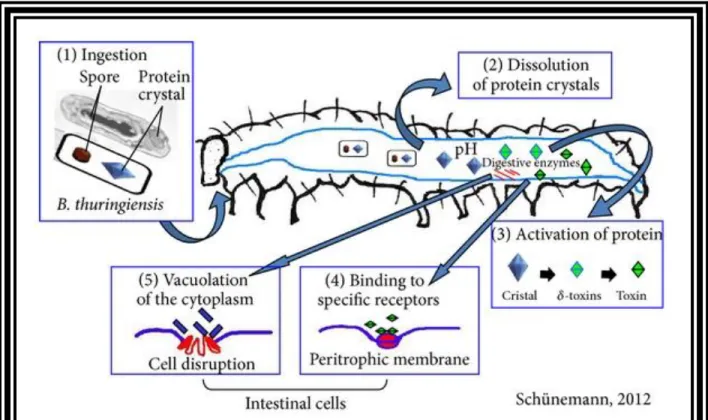


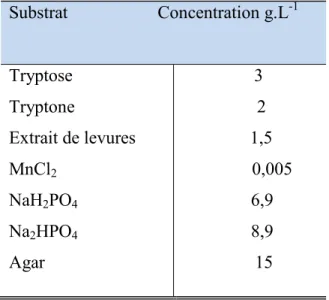

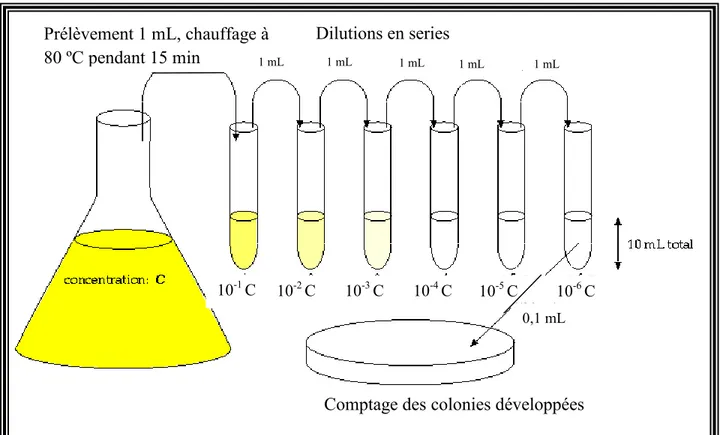
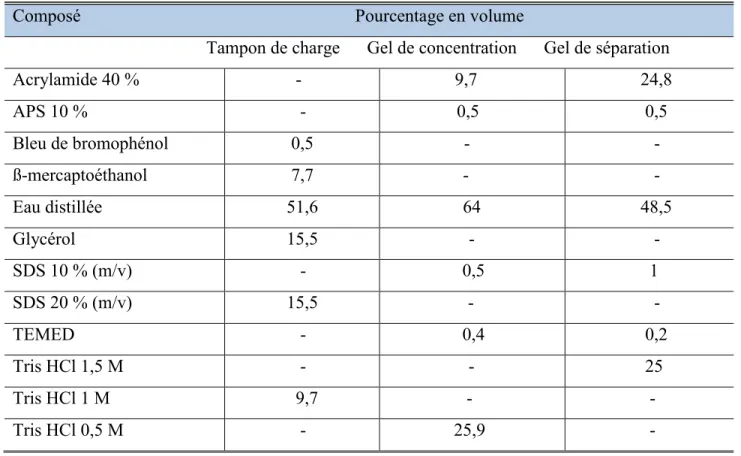


Documents relatifs
Un autre milieu solide sélectif permettant de tester le caractère acidifiant des levures présentes d'une part et leur aptitude à convertir l'acide p-coumarique en 4-éthylphénol
L'effet de différentes concentrations de fer dans le milieu de culture sur la production de rhamnolipides a été comparé. Ces paramètres ont été suivis: la croissance,
La présence d’un facteur inhibiteur de la ribonucléase dans le milieu réactionnel entraîne une diminution de la quantité d’oligonucléotides libérés, donc
Les membres du groupe bacillus cereus sont des contaminants fréquents de l'environnement laitier, et sont largement impliqués dans les problémes de contamination et de dégradation
En définitive, une corrélation entre les concentrations dans les moules et celles dans I'eau peut seulement être attendue dans le cas où lEvolution de la contamination du milieu
Dans ce milieu, les éponges marines sont largement étudiées car elles abritent de nombreux micro-organismes différents (Li et al., 2006) et produisant des
Lors de l’ajout d’AMG 0.05X dans la formulation de l’hydrogel, une production plus faible de glucose est observée (20% de la quantité maximale de glucose qui peut être produit
A pH 7,4 dans le milieu des cellules proliférantes ou dans le milieu des cellules quiescentes, la molécule a un effet faible sur les cellules en quiescence (plateau de
